Two different enzymes condition the sucrose synthase activity that is detected in the maize plant (Chourey, MGG 184:372, 1981; Echt & Chourey, Plant Physiol. 79:530, 1985). Genes coding for these enzymic activities have been identified, cloned, sequenced and genetically mapped (Chourey & Nelson, Biochem. Genet. 14:1041, 1976; Geiser et al., EMBO J. 1: 1455, 1982; Werr et al., EMBO J. 4:1373, 1985; Sheldon et al., MGG 190:421, 1983; Zack et al., Maydica 31:5, 1986; McCarty et al., PNAS 83:9099, 1986; Shaw and Hannah, unpublished). Whereas the activity coded for by the Sh locus is found almost exclusively in the endosperm, the activity coded for by the second gene is found in the endosperm, embryo, roots, shoots and cultured cells (see references above). To our knowledge, there is only 1 report which describes a maize tissue lacking sucrose synthase activity. W.H. Bryce and O.E. Nelson (Plant Physiol. 63:318, 1979) reported that pollen of maize lacked this enzymic activity whereas detectable levels of several other starch synthetic enzymes were found in this tissue.
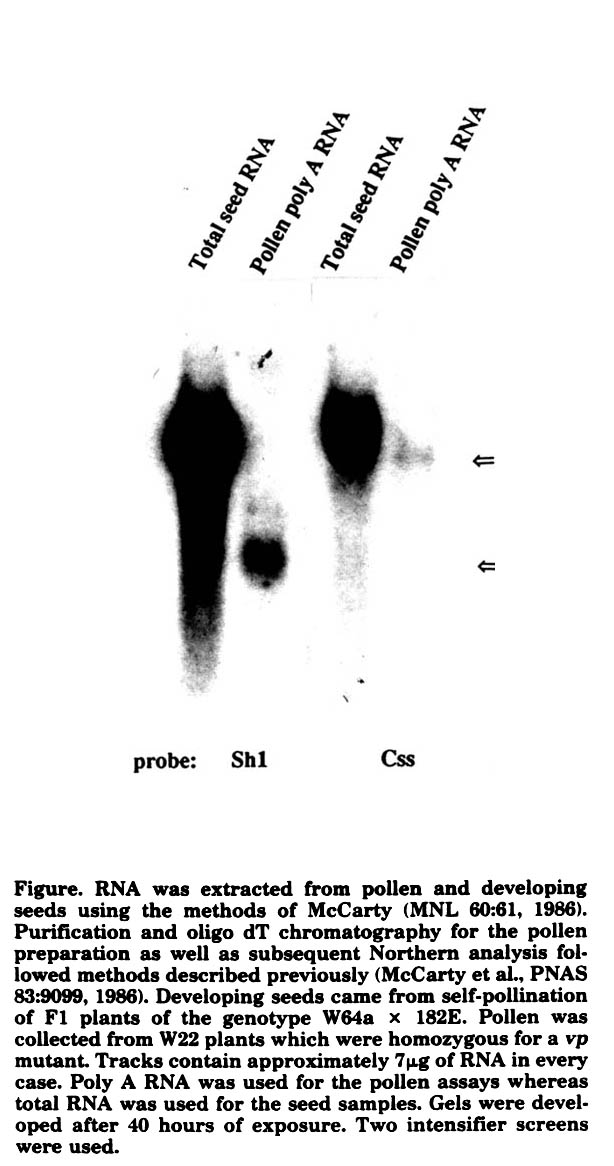
To ask more directly whether either of the 2 structural genes for sucrose synthase is expressed in the maize pollen, Northern analyses of RNA from the pollen and from seeds were performed. As shown in the figure, Css MRNA having the same mobility as that of the seed is detected in the mature pollen of maize. Thus it would appear that the Css locus is expressed in this tissue as well as in most if not all tissues of the maize plant.
Perhaps unexpectedly, the Sh probe also detects a transcript in the mature maize pollen. The transcript is significantly smaller than the Sh transcript of the endosperm. The origin and genetic control of this mRNA are under investigation.
Chourey (MNL 61:55, 1987) has correctly pointed out some of the confusion associated with the nomenclature of the genes encoding sucrose synthase enzymes of maize. The major problem arises from the fact that the gene shrunken, or Sh, which encodes the predominant endosperm sucrose synthase was named for its characteristic kernel phenotype and not for its biochemical function. Because of this, the naming of the gene coding for the second sucrose synthase has not been trivial.
We (McCarty et al., MNL 60:58, 1986) had initially used the abbreviation Ss2 as the gene symbol for the second gene. However, three problems with this symbolism became apparent during the review of the paper describing this gene. The criticisms are the following: (1) Ss2 is misleading in that there is not an Ss1 or Ss. In order to have an Ss, Sh would have to be changed to Ss or we would have to have 2 symbols for 1 gene. Neither possibility is particularly appealing to us. (2) If in fact Ss is really Sh then, the argument goes, Ss2 must really be Sh2. This, of course, is wrong. Sh2 is on another chromosome and affects a step later in the starch biosynthetic path, ADP-glucose pyrophosphorylase. Although we initially thought that this was common knowledge, the mistake of equating Ss2 with Sh2 has been made on many occasions (public talks, question and answer sessions, poster sessions and even in our own lab meetings). Continued usage of Ss2 and the compelling logic in the above argument will quickly lead to 1 gene symbol for 2 genes. (3) Ss2 does not follow the recommendations of our group concerning gene symbols. One need only look to recommendation 1 (Burnham et al., MNL 49:3, 1975) to see that all new genes (after 1975) should be given a three letter symbol. This point clearly rules out Ss2 if we are to follow our own rules. As an alternative, we have used Css for constitutive sucrose synthase. The symbol has a number of attractive features: (1) It gives a fair description of the expression of the gene. While we have not examined every cell in every tissue of a corn plant, the data to date point to the fact that Css is expressed in many if not all tissues of the plant. As we have pointed out above, Css transcripts can be found in a tissue which reportedly lacks sucrose synthase activity. Whether this reflects a form of control at the translational level or differences in sensitivity of the enzyme assay versus RNA assay is presently unknown; however, there is now no reason to think that the gene is not expressed in this tissue. The term constitutive should not be equated with non-inducible. An examination of the literature concerning the original operator-constitutiue mutants of the lac operon of E. coli shows that these mutants were still inducible; the level of induction, however, is less than that of wild type. (2) It is difficult to confuse Css with Sh2. The symbol Css, like Sh1, differs substantially from the words sucrose synthase. (3) Css follows the rules of maize nomenclature.
It is our hope that the problems of gene symbolism surrounding the sucrose synthases can now be resolved.
Figure. RNA was extracted from pollen and developing seeds using the methods of McCarty (MNL 60:61, 1986). Purification and oligo dT chromatography for the pollen preparation as well as subsequent Northern analysis followed methods described previously (McCarty et al., PNAS 83:9099, 1986). Developing seeds came from self-pollination of F1 plants of the genotype W64a X 182E. Pollen was collected from W22 plants which were homozygous for a vp mutant. Tracks contain approximately 7µg of RNA in every case. Poly A RNA was used for the pollen assays whereas total RNA was used for the seed samples. Gels were developed after 40 hours of exposure. Two intensifier screens were used.
L.C. Hannah and D.R. McCarty
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}