The ability to analyze the functions of the Suppressor-mutator (Spm) element by in vitro mutagenesis techniques requires that the element be able to function in a transformable cell system. It has been established that the Activator element transposes at a high frequency in tobacco cells when introduced by Agrobacterium-mediated transformation (Baker et al., Proc. Natl. Acad. Sci. 83:4844, 1986; EMBO J. 6:1547, 1987). We have obtained preliminary evidence that the Spm element also moves in tobacco cells.
To test the ability of the Spm element to both excise and trans-activate excision in tobacco, we constructed derivatives of the binary Agrobacterium vector pMON410, developed by Monsanto Co. (Rogers et al., Meth. Enzymol. 118:627, 1986). It carries 2 bacterial drug-resistance marker genes, both of which have been modified for expression in plant cells by the substitution of a plant promoter and the addition of a plant polyadenylation sequence. The plasmid is capable of conferring resistance to both kanamycin and hygromycin on transformed tobacco cells.
The pMON410 plasmid was modified by the introduction of a transposition-defective Spm (dSpm) element between the promoter and the hygromycin resistance marker (Fig. 1). The dSpm element was constructed by deleting the element's 2 internal EcoRV restriction fragments (Pereira et al., EMBO J. 5:835, 1986; Masson et al., Genetics 177:117, 1987). In addition, a complete Spm element was inserted at a unique cloning site near the plasmid's kanamycin-resistance marker (neomycin phosphotransferase II. The relevant parts of the plasmid's structure are shown in Figure 1.
The modified plasmid containing both the dSpm and Spm elements, and control plasmids lacking one or both Spm insertions, were introduced into tobacco cells by the leaf disc transformation technique (Rogers et al., 1986). Transformation was judged by the development of kanamycin resistant calli as a consequence of the introduction of the MPTII gene of the pMON410 plasmid. The ability of the plasmid-borne Spm elements to excise was assessed both by direct analysis of the T-DNA structure of transformed cells, and by the acquisition of hygromycin resistance by transformed cells.
Because the dSpm element is inserted at a site between its promoter and the hygromycin-resistance marker, the modified plasmid should not be capable of conferring hygromycin resistance on transformed cells. Control experiments with a plasmid containing the dSpm element but no Spm element established that the dSpm insertion disrupts expression of the hygromycin-resistance gene. If an Spm element residing on the plasmid can catalyze excision of the dSpm element, then transformed cells in which such an excision event has occurred will be resistant to both kanamycin and hygromycin.
Selection for hygromycin resistance has been applied both immediately after transformation and after the development of calli on kanamycin-containing medium. The number of hygromycin-resistant calli was the same for the experimental plasmid and the control plasmid lacking the trans-acting Spm element when hygromycin selection was applied immediately after transformation. Hygromycin resistance has been observed in calli initially selected for kanamycin resistance, but the structure of the resident T-DNA has not yet been analyzed.
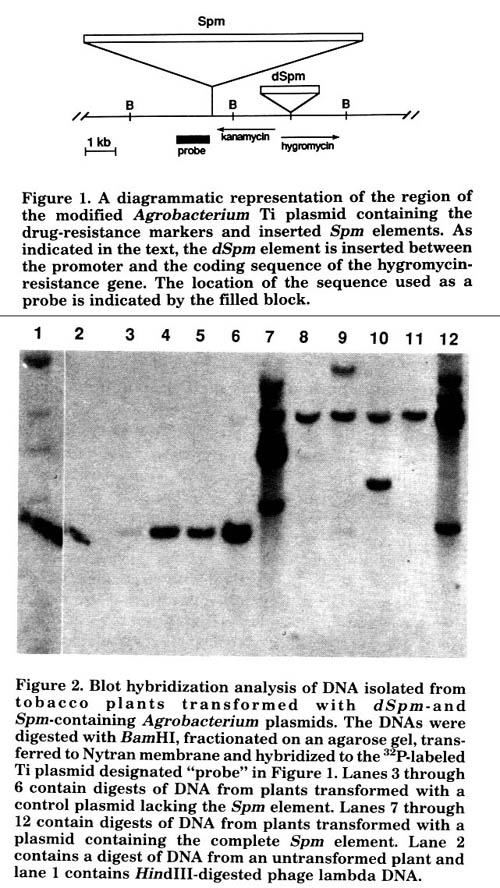
However, the resident T-DNA has been analyzed in DNA extracted from several plants regenerated from kanamycin-resistant calli transformed with Agrobacteria carrying experimental and control plasmids. Different probes of T-DNA structure were used to determine whether the intact element and the dSpm element were moving. The results of one such experiment are shown in Figure 2. The DNA was digested with BamHI and probed with a fragment whose location is shown in Figure 1 to determine whether the complete Spm element was present on the plasmid.
All of the DNAs from calli transformed with a plasmid lacking the Spm (lanes 3-6) exhibited only a 3.5-kb BamHI fragment with homology to the probe. This represents the undisrupted Spm insertion site. All of the DNAs from calli transformed with a plasmid containing the Spm element exhibit the expected 8.5kb fragment comprising part of the Spm element and the plasmid sequence flanking it in the original construct. In addition, the DNA samples in lanes 9 and 12 show a fragment with homology to the probe that is about 3.6kb in length. The DNA sample in lane 11 also showed a fragment of the same mobility after a longer exposure. The 3.6kb fragment is likely to represent an empty donor site from which the element has excised, since the inserted element was flanked by about 0.1kb of sequence from the maize a locus from which it was originally cloned.
Additional evidence that excision is occurring has been provided by comparable results of analyses of the dSpm insertion site (not shown). Not surprisingly, the same DNA samples that exhibit excision of the Spm element give evidence of dSpm excision. Taken together, these observations provide encouraging preliminary evidence that the Spm element is mobile in tobacco. Experiments are under way to clone Spm-containing sequences from tobacco to provide direct evidence for excision and transposition.
Figure 1. A diagrammatic representation of the region of the modified Agrobacterium Ti plasmid containing the drug-resistance markers and inserted Spm elements. As indicated in the text, the dSpm element is inserted between the promoter and the coding sequence of the hygromycin-resistance gene. The location of the sequence used as a probe is indicated by the filled block.
Figure 2. Blot hybridization analysis of DNA isolated from tobacco plants transformed with dSpm-and Spm-containing Agrobacterium plasmids. The DNAs were digested with BamHI, fractionated on an agarose gel, transferred to Nytran membrane and hybridized to the 32P-labeled Ti plasmid designated "probe" in Figure 1. Lanes 3 through 6 contain digests of DNA from plants transformed with a control plasmid lacking the Spm element Lanes 7 through 12 contain digests of DNA from plants transformed with a plasmid containing the complete Spm element. Lane 2 contains a digest of DNA from an untransformed plant and lane 1 contains HindIII-digested phage lambda DNA.
Patrick Masson, Kathy Toohey and Nina Fedoroff
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}