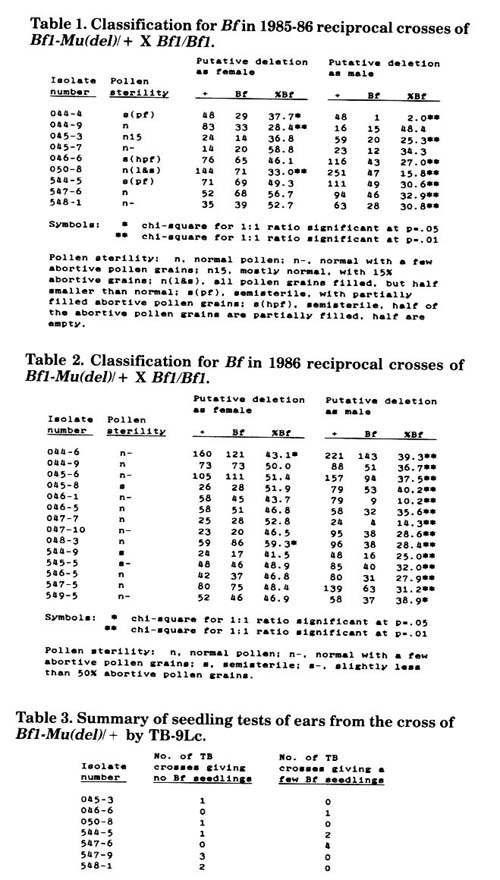
In 1986, we reported preliminary evidence suggesting that deletions had been induced by Mutator that include the Bf1 locus (MNL 60:11-12). The original isolates carrying the putative deletions were of the genotype Bf1/Bf1 - Mu(del). All isolates were crossed by standard lines (+ / +). In our 1985-86 winter nursery, kernels from 9 of these crosses of the putative deletion isolates by standard were planted, and the resulting plants reciprocally crossed with homozygous Bf1 testers; the progeny of these reciprocal crosses were seedling tested and scored for Bf1 (Table 1). Seven of the isolates had the pattern of segregation expected if a deletion were present (i.e., a deficiency of Bf1 transmission in male outcrosses of putative deletion heterozygotes to Bf1 testers). Isolates 044-9 and 045-7 showed uncertain indication of a deletion being present; isolate 044-9 showed some reduced female transmission of Bf1 but normal male transmission, and isolate 045-7 did not produce a statistically significant sample. Two isolates showed reduced transmission of Bf1 through both the male and female.
In our 1986 summer nursery, reciprocal crosses to Bf1 testers were made of 19 additional putative deletion isolates that had been crossed by standard. Isolate 044-9, which gave ambiguous results in Table 1, was retested as well. Fourteen of these showed reduced male transmission of Bf1 indicative of the presence of a deletion (Table 2); of these 14, one showed reduced female transmission as well. Among this set of isolates, as well as those presented in Table 1, there did not seem to be a consistent association of pollen sterility with reduced male transmission. Thus, pollen sterility does not seem to be invariably associated with deletions involving Bf1.
In 1986, kernels from 8 crosses of putative deletion isolates by standard were planted and the resulting plants pollinated by TB-9Lc pollen. Each plant was also outcrossed to a Bf1 tester to determine which ones carried the putative deletions. Approximately 50 kernels from each ear crossed by TB were sown in the seedling bench and scored for Bf1. Some of the TB crosses of plants heterozygous for a putative deletion segregated for a few Bf seedlings, and some did not (Table 3). Thus, if deletions are indeed present in all of these stocks, it seems that some deletions are viable in the hemizygous condition (i.e., those in which Bf seedlings were found), while others are not (i.e., those in which no Bf seedlings were found). However, more seedlings of the latter class should be tested before it can be said with certainty that these putative deletions will not survive in the hemizygous state.
Last summer, Bf seedlings from the TB crosses were transplanted into the field. Following are the putative deletions tested and, in parentheses, the number of plants observed for each: 046-6 (4), 544-5 (3), 547-6 (12), and 548-1 (5). In all cases, these plants had tassels that extruded few, if any, anthers. They also were later maturing, were reduced in height, and had narrow leaves. The latter 3 traits are characteristic of hypoploid TB-9Lc plants, but the lack of anther extrusion is not. It would seem that if deletions are indeed present, these deletions are missing loci necessary for anther extrusion. It was also of interest to note that all plants had green midribs. If deletions are present that include Bf1, they seemingly do not extend to the bm4 locus, which is distal to Bf1. Thus, these would not seem to be terminal deletions. We are currently growing test crosses of our Bf1 deletion lines by bm4 to verify whether this is so.
Dr. Earl Patterson has recently located at least one male sterile mutant to the long arm of chromosome 9 by crossing with TB-9Lc. Crosses of this male sterile by our putative Bf1 deletion stocks that have reduced male transmission will be made to determine whether this male sterile locus is uncovered by our deletions.
In summary, there is genetic evidence of Mutator having induced deletions which include the Bf1 locus on the long arm of chromosome 9, some of which are lethal in the hemizygous condition, and at least some of which do not include the bm4 locus and are thus not terminal. Those deletions tested to date also seem to involve a locus controlling anther extrusion.
Table 1. Classification for Bf in 1985-86 reciprocal crosses of Bf1-Mu(del)/ + X Bf1/Bf1.
Table 2. Classification for Bf in 1986 reciprocal crosses of Bf1-Mu(del)/ + X Bf1/Bf1.
Table 3. Summary of seedling tests of ears from the cross of Bf1-Mu(del)/ + by TB-9Lc.
Donald S. Robertson and Philip S. Stinard
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}