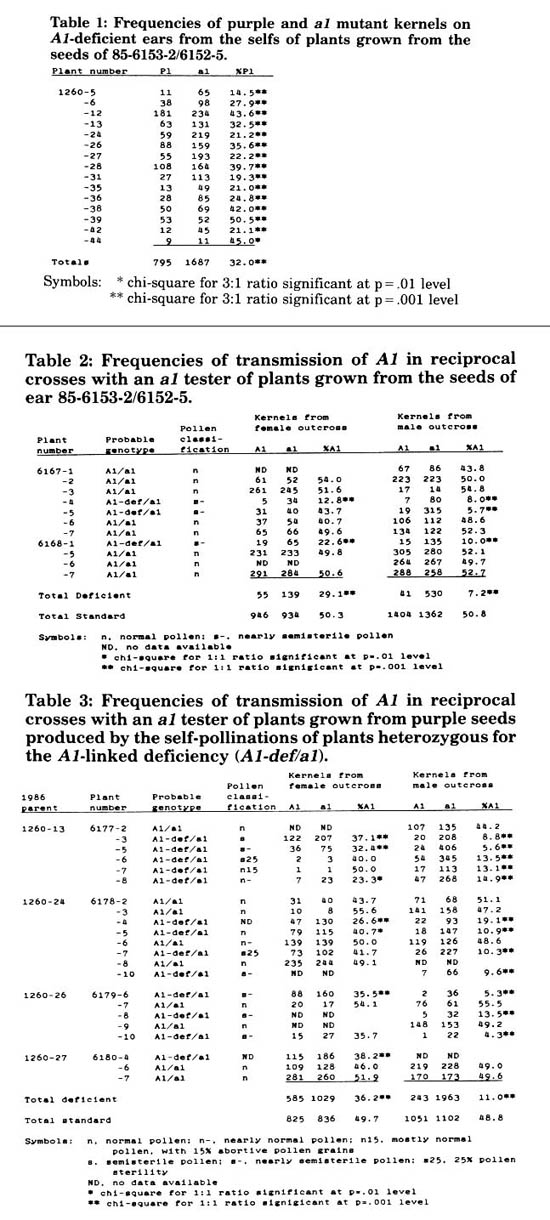
In our 1985 summer nursery, we crossed a purple aleurone Mutator (Pl Mu) plant by a Mutator-induced a1 mutable stock (a1-Mum3) as part of our ongoing Mutator inbreeding experiments. Kernels from this cross (85-6153-2/6152-5) were planted in our 1986 selfing block, and the resulting plants self-pollinated. Of 44 selfs, 15 ears segregated for ratios of purple to mutant kernels that deviated greatly from the expected 3:1 ratio. In each instance, the purple class was deficient, resulting in ratios as low as 1:4 (Table 1). These same ears were also semisterile. We hypothesized that the low frequency of transmission of A1 in these selfs was due to a deficiency linked to A1 that is not transmissible through the male, and probably only poorly transmitted through the female. The purple kernels appearing on the deficient selfed ears were either the result of crossovers with the chromosome 3 homologue not carrying the deficiency, or the result of reduced female transmission of the deficiency. The deficiency arose in the Pl Mu parent (or its progenitor) on one of the chromosome 3 homologues, and was transmitted to its progeny in a reduced frequency (15/44). In order to study this phenomenon further, we planted more kernels from 85-6153-2/6152-5 in our 1987 crossing block, and reciprocally crossed the resulting plants with a1 testers. We also planted purple kernels from the deficient selfed ears, and reciprocally crossed the resulting plants with a1 testers.
Of 11 plants grown from the kernels of 85-6153-2/6152-5, only 3 carried the deficiency (Table 2). Male transmission of the A1 allele in reciprocal crosses with a1 testers was greatly reduced (7.2%), but female transmission was reduced as well (29.1%). Pollen shed from anthers collected from deficient plants was semisterile, probably reflecting gametophytic lethality associated with pollen carrying the deficiency.
Of 22 plants grown from the purple kernels of deficient selfed ears, 13 carried the A1-linked deficiency (Table 3). Again, both male and female transmission of the A1 allele was reduced in reciprocal crosses with a1 testers (11.0% and 36.2% respectively), and pollen produced by these plants was semisterile (with the exception of one which had only 15% abortive pollen). The 9 plants from this set that did not carry the deficiency (as evidenced by the equal transmission of A1 and a1 through both the male and the female) probably reflect crossovers occurring in the parent during either mega- or microsporogenesis that placed A1 in coupling with non-deficient chromosome 3 segments.
We made crosses of deficient plants by homozygous etched stocks, but recovered no etched kernels, indicating that the deficiency does not include the etched locus.
It is possible to derive both a percent recombination of A1 with the deficiency, and a rate of female transmission of the deficiency, if one makes the following assumptions: (1) the deficiency is not male transmissible; (2) the percentage recombination between A1 and the deficiency is the same in both the male and the female; (3) the deficiency is transmitted through the female with the same frequency in both the parental and crossover classes. With these assumptions, and combining the data for the reciprocal crosses of deficient plants in Tables 2 and 3, one arrives at a percent recombination of A1 with the deficiency of 10.2 ± 0.6, and a female transmission rate of 31.7% (i.e. in a plant heterozygous for the deficiency, 31.7% of the ovules will carry the deletion). This female transmission frequency agrees well with the percentage of the offspring of 85-6153-2/6152-5 that carry the deficiency (18/55 = 32.7%). One can use the calculated percent recombination and female transmission frequency to deduce expected frequencies of purple and mutant kernels in selfed ears of deficiency heterozygotes. The expected frequency of purple kernels in such selfs, 42.0%, differs significantly from the average frequency of purple kernels observed in the selfed plants from Table 1 (32.0%), but comes close to several individual values.
We plan to conduct further experiments with other linkage markers to determine whether the deficiency is proximal or distal to A1. We will also try to recover the crossover class that places the deficiency in coupling with a1. If this deficiency is Mutator-induced, it should also be amenable to sibling segregation analysis to determine whether a Mutator element co-segregates with the deficiency.
Table 1: Frequencies of purple and a1 mutant kernels on A1-deficient ears from the selfs of plants grown from the seeds of 85-6153-2/6152-5.
Table 2: Frequencies of transmission of A1 in reciprocal crosses with an a1 tester of plants grown from the seeds of ear 85-6153-2/6152-5.
Table 3: Frequencies of transmission of A1 in reciprocal crosses with an a1 tester of plants grown from purple seeds produced by the self-pollinations of plants heterozygous for the A1-linked deficiency (A1-def/a1).
Philip S. Stinard and Donald S. Robertson
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}