Evidence has accumulated that deletions have been induced by Mutator at the yg2 and Bf1 gene loci (see subsequent articles in this News Letter). We now present evidence of a Mutator-induced deletion of a segment of chromosome 3 including the A1 and Sh2 loci.
In our 1985-86 winter nursery, we observed a 3-seed sector of yellow brittle kernels on an ear (85-86-6512-6/5513-3) from the cross of a purple aleurone Mutator stock by an a1-Mum2 Sh2 stock heterozygous for the standard a1 sh2 tester alleles in coupling. Since our purple Mutator stocks are homozygous for the A1 and Sh2 alleles, we hypothesize that the 3-seed sector arose premeiotically by the deletion of a small segment including the A1 and Sh2 loci (located 0.2 cM apart) on one of the chromosome 3 homologues of the purple Mutator parent. The eggs arising from the ear sector carrying this deletion were fertilized by a1 sh2 pollen from the male parent, giving rise to the sector of 3 yellow brittle seeds. Since the male parent was heterozygous for a1 sh2, there is a possibility that some eggs carrying the deletion could have been fertilized by pollen carrying the a1-Mum2 and Sh2 alleles. Kernels arising from such a fertilization would be starchy, and either mutable or yellow stable. No such kernels were observed.
In order to analyze this putative deletion genetically, we attempted to germinate the 3 yellow brittle kernels in Petri dishes. Two of the 3 kernels molded, but the third kernel produced a small, weak seedling that grew slowly and reached silking stage about 90 days after germination. We had intended to pollinate this plant (86-8501-1) by a standard purple aleurone stock in order to separate the standard a1 sh2 homologue from the a1 sh2 deletion homologue, but instead pollinated by c1 c1 r r B79 inbreds (86-908-4 and 86-910-1) on two successive days, as these were the only plants shedding pollen late in the season. This pollination accomplished the same result, as B79 is wildtype for A1 and Sh2. The ear resulting from this cross (86-8501-1/908-4 & 910-1) had normal seed set and produced about 100 kernels. We would expect about half of these kernels to be A1 Sh2/a1 sh2-deletion, and half to be A1 Sh2/a1 sh2-standard.
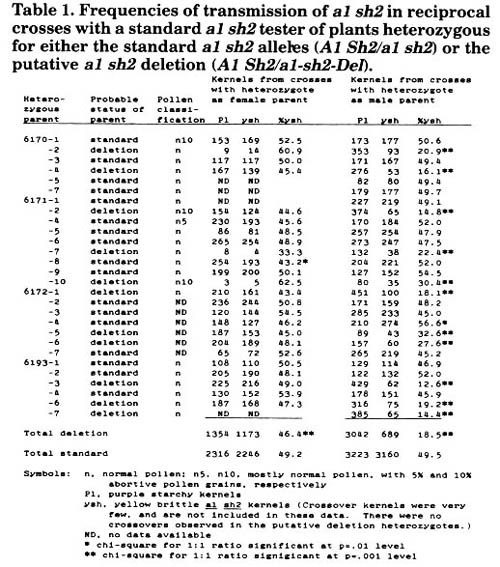
Kernels from 86-8501-1/908-4 & 910-1 were grown in our 1987 summer nursery, and the resulting plants were reciprocally crossed with a homozygous standard a1 sh2 tester on the first ear, and pollinated by either a purple aleurone stock or an a1 a1 et et stock on the second ear. In addition, shedding tassel branches were sampled from as many of these plants as possible, and the pollen examined for sterility. The results of the reciprocal crosses with standard a1 sh2 are presented in Table 1. Out of 27 pairs
of reciprocals made, 11 were significantly deficient (p < 0.001) in the yellow brittle class when the heterozygous parent was outcrossed as a male. These 11 heterozygotes presumably carry the putative a1 sh2 deletion, and transmit the deletion in a highly reduced frequency when the deletion pollen competes with wildtype pollen. The average frequency of transmission of a1 sh2 through the male in these 11 plants is 18.5%, but the individual frequencies are very heterogeneous (homogeneity chi-square p < 0.001). The putative deletion seems to be fully transmissible through the female if one looks at individual ears, since none of the crosses of the putative deletion heterozygotes as females by standard a1 sh2 gave rise to ears deviating significantly from 1:1 in the ratio of purple starchy to yellow brittle kernels. However, when one looks at the totals for the deficient ears, the average percent transmission of a1 sh2 through the female is 46.4%, significantly different from a 1:1 ratio (p < 0.001). Thus, female transmission of the putative deletion may be very slightly reduced. The pollen samples from all putative deletion heterozygotes examined were normal, or near normal, with no evidence of semisterility. Apparently, the major effect of the deletion on the pollen is to reduce its competitive ability with wildtype pollen. Whether this is accomplished by reduced pollen germination, by slower pollen tube growth, or some other mechanism, is not known. We have examined the pollen from a known a1 sh2 deletion heterozygote (a1-x1) provided by the Maize Coop, and find that it is semisterile. Perhaps the a1-x1 deletion includes regions essential for the growth of the male gametophyte that are not deleted in our putative a1 sh2 deletion. Cytological analysis may be able to help verify whether this is true.
Crosses of putative deletion heterozygotes by etched stocks gave rise to only non-etched kernels. Thus, the putative deletion does not include the etched locus.
We have not analyzed the putative deletion molecularly, but we hope that this may soon be accomplished by either ourselves or others. Should this prove to be a deletion, and should a copy of a Mutator element be found at the site of the deletion, this would shed further light on the role of Mutator in inducing deletions. Furthermore, since we have sibling kernels from the ear on which the putative deletion was observed, one could investigate molecularly the possible mechanisms for the formation of the deletion (e.g. pre-meiotic unequal sister chromatid exchange or somatic crossing over, occurring by slant pairing of homologous Mutator elements on either side of the A1 and Sh2 loci).
Table 1. Frequencies of transmission of a1 sh2 in reciprocal crosses with a standard a1 sh2 tester of plants heterozygous for either the standard a] sh2 alleles (A1 Sh2/a1 sh2) or the putative a1 sh2 deletion (A1 Sh2/a1-sh2-Del).
Philip S. Stinard and Donald S. Robertson
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}