Tissue culture allows an important extension of studies of certain in planta phenomena. These methods have been used in a limited way to analyze the activity of Activator and Enhancer transposable element systems in maize endosperm cultures (A.R. Reddy and P.A. Peterson, Maydica 22:125, 1977; Culley, VI International Congress Plant Tissue Culture p. 220, 1986), and more extensively in the analysis of transgenic expression of Ac activity in tobacco cells (B. Baker et al., Proc. Natl. Acad. Sci. USA 83:4844, 1986). Research to date on the Mutator system has centered primarily upon the analysis of differentiated maize tissues in planta, including leaves, seedlings, inmature ears, and kernels, although endosperm cultures derived from lines with Mu-element-containing mutable alleles have recently been established in this laboratory. Endosperm callus, however, grows relatively slowly, making it difficult to accumulate quantities of material sufficient for extensive molecular study. In contrast, Type II embryogenic callus lines of maize (C.L. Armstrong and C.E. Green, Planta 164:207,1985) are reliably fast-growing and allow for continued analysis of an expandable pool of genotypically identical tissue as well as the possibility of plant regeneration. To study Mutator systems in tissue culture we have, therefore, established Type II callus lines from immature F1 embryos of crosses of mutagenically active Mutator stock (Mu2) with inbred lines A188 and H99.
We report our initial molecular analysis of Mu-elements in 13 independently derived embryogenic callus cultures. Active Mutator lines of maize in planta are characterized by their ability to generate new mutants at 30 to 50 times the spontaneous level, and by somatic instability at Mutator-induced mutant alleles. To date, mutagenically active Mutator lines with fewer than 10 Mu-elements per diploid genome have not been observed (J.L. Bennetzen et al., Mol. Gen. Genet. 208:45, 1987), and stable maintenance of a high Mu-element copy number from one generation to another is thought to be a feature of an active Mutator system (V. Walbot and Warren, Mol. Gen. Genet., in press, 1987). Mutator activity also correlates with lack of modification at the HinfI restriction sites which lie within the inverted terminal repeats of Mu-elements (V.L. Chandler and V. Walbot, Proc. Natl. Acad. Sci. USA 83:1767, 1986). Because of the correlation of these parameters with Mutator activity, we have investigated Mu copy number and HinfI-site modification at both early and late times in culture. Mu modification was assessed by genomic digests with the methylation sensitive enzyme HinfI (S. Hake and V. Walbot, Chromosoma 79:251, 1980), which recognizes sites in the terminal inverted repeats of this element. Subsequent hybridization of unmodified transposons with a Mu1 probe results in discrete 1.3 and 1.6 kb bands which represent the Mu1 and Mu1.7 elements, respectively. However, similar treatment of DNA which contains Mu-elements modified at Hinfl sites results in a ladder of bands larger than 1.3 kb. Mu-element copy number was determined by comparison with plasmid copy-number reconstructions or by analysis of the genomic location of all elements present.
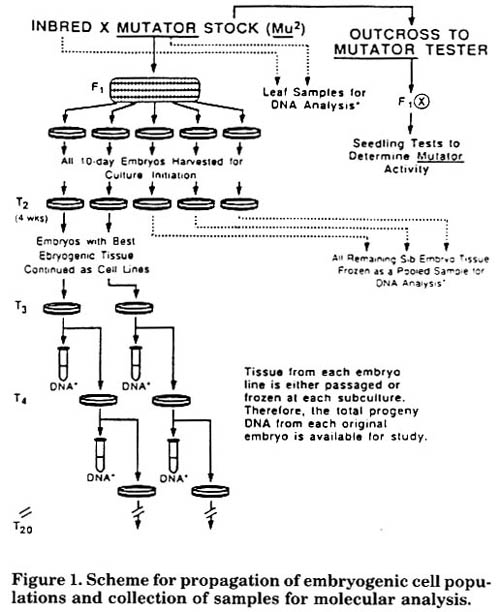
All progeny embryos from each inbred/Mutator F1 ear were cultured. Four and 6 weeks after initiation it was possible to identify individual embryos with good Type II embryogenic callus. To establish individual embryo lines these were selected for transfer to fresh medium, and all other calli and tissues were pooled and frozen at -20 C. Thereafter, in each individual line Type II callus with the best embryogenic phenotype was transferred and propagated, and all nonselected tissues were stored separately at -20 C for subsequent molecular analysis. In this manner, all tissue descending from each individual embryo population was saved in the fresh or frozen state (Fig. 1). DNA isolated from sib embryos pooled at 4 and 6 weeks (T2 and T3) was analyzed molecularly to enable the earliest possible characterization of Mu elements in an embryogenic callus culture.
To determine the effects of culturing on Mu-element copy number and modification state, it is necessary to make comparisons with parental plant controls. Leaf samples taken at the 4- to 7-leaf stage from Mutator parents were used for molecular analysis of Mu-element copy number and HinfI-site modification. In addition, Mu2 parent plants were tested genetically for Mutator activity (D.S. Robertson, Mol. Gen. Genet. 183:51, 1981).
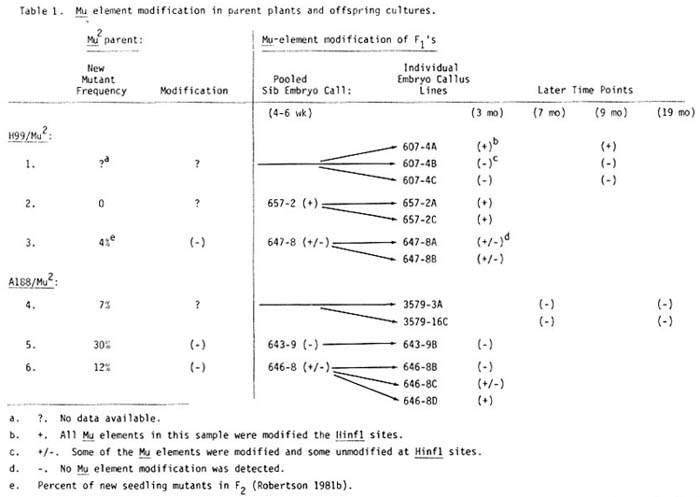
Because plant progeny which result from outcrosses of active Mutator stock have Mu elements which are unmodified at the HinfI sites (J.L. Bennetzen, Mol. Gen. Genet. 208:45, 1987), it was expected that embryogenic cultures established from the F1 progeny of an outcross between an active Mutator line with A188 or H99 would have similarly unmodified Mu elements. Our results show, however, that the state of Mu-element modification varies in the F1 cultures examined even though the Mu2 parent of the culture was mutagenically active (Table 1, lines 3, 4, 5, and 6). Among the six A188-derived embryogenic lines examined, one culture has Mu elements uniformly modified at the HinfI sites, four have Mu elements which are unmodified at these sites, and one has a population of Mu elements with a mixed state of modification. A similar variation occurs when H99 is the inbred parent. Three of the 7 H99/Mu2 F1 cultures have Mu elements which are uniformly modified at the HinfI sites, 2 have only Mu elements which are unmodified at these sites, and 2 have both modified and unmodified elements. It should be noted, however, that one Mutator parent of 2 HinfI -modified cultures from H99 was not mutagenically active (Table 1, line 2). Control callus cultures, generated by crossing H99 with inactive Mutator parents (Mu16 and Mu32) show the complete Mu element-HinfI site modification expected in the outcross progeny of Mutator lines previously made inactive by repeated intercrossing.
Comparisons of the molecular profiles of early pooled samples of sib embryos with related F1 embryo lines show that the Mu-modification state in the individual lines was always like that seen in the early pooled samples (Table 1). This indicates that the unexpected modification of Mu-element HinfI sites, when seen, was an event which probably occurred early in culture, before 4 weeks.
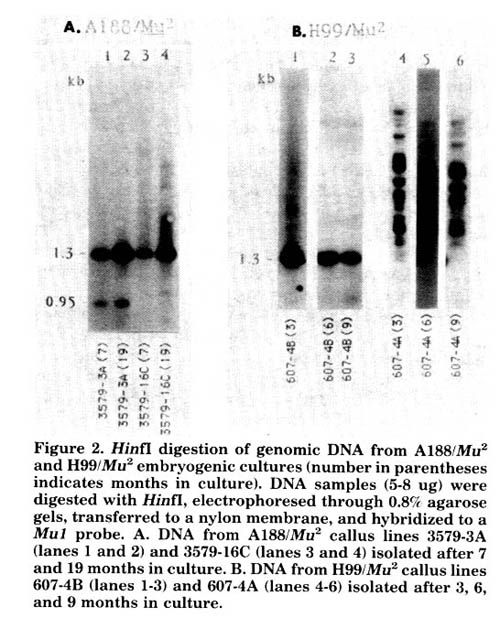
Two A188/Mutator callus lines have been in continuous culture for almost 2 years (3579-3A and 3579-16C, Fig. 2). Analysis of the DNA isolated from tissue harvested after 7 and 19 months in culture shows no modification at the Mu-element HinfI sites in either line. Two sibling H99/Mu2 lines were also examined molecularly after 3, 6, and 9 months in culture (607-4A and 607-4B, Fig. 2). One line was completely and stably modified at the Mu-element HinfI sites over this time period, while the other was stably unmodified. The basic stability of the state of Hinfl-site modification in each line examined suggests that continued culturing does not reverse modification patterns that are established early in culture.
The fact that Mu elements in approximately half of the cultures examined did exhibit HinfI site modification is surprising and suggests that it may be associated with the establishment of the maize embryos in culture. Culturing of plant tissues has been shown to result in increased variability (Larkin and Scowcroft, Theor. Appl. Genet. 60:197, 1981), with generalized changes in the methylation of genomic DNA thought to be due to tissue culture stress (Brown and Lorz, in Somaclonal Variations in Crop Improvement, 1986). Additional analysis of these cultures with other methylation-sensitive restriction enzymes and probing with developmentally active and inactive genes will be used to test the specificity of the modification seen.
All callus lines studied contain approximately 15 to 25 Mu elements and these are also stably retained. Lines which have unmodified Mu elements in this high copy number are potentially active Mutator systems. Transposition of Mu elements in these cultures is being tested in experiments designed to fix such genomic events in callus-line subpopulations. Our laboratory has also established embryogenic cultures from plant lines which have Mu elements at specific mutable alleles (wx, bz1, and a1), and Mu element excision from these loci in culture will be investigated. These studies should further determine the potential usefulness of the Mutator system in culture.
Table 1. Mu element modification in parent plants and offspring cultures.
Figure 1. Scheme for propagation of embryogenic cell populations and collection of samples for molecular analysis.
Figure 2. HinfI digestion of genomic DNA from A188/Mu2 and H99/Mu2 embryogenic cultures (number in parentheses indicates months in culture). DNA samples (5-8 ug) were digested with HinfI, electrophoresed through 0.8% agarose gels, transferred to a nylon membrane, and hybridized to a Mu1 probe. A. DNA from A188/Mu2 callus lines 3579-3A (lanes 1 and 2) and 3579-16C (lanes 3 and 4) isolated after 7 and 19 months in culture. B. DNA from H99/Mu2 callus lines 607-4B (lanes 1-3) and 607-4A (lanes 4-6) isolated after 3, 6, and 9 months in culture.
Martha James and J. Stadler
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}