The lack of an effective transformation system to introduce foreign and/or engineered DNA into developing, reproductively competent maize cells continues to be one of the major limitations in plant molecular genetics. One approach to the delivery of exogenous DNA into maize involves the direct transformation of gametes. This technology could circumvent the expense, time, and genome stress associated with the transformation of cells in culture and subsequent plant regeneration. The rapid cell division and high level of cell-cell communication in the developing maize tassel provides a possible window of opportunity for the introduction of DNA into male gametocytes. Upon cloning the Adh1 locus in 1982, we decided to macroinject this gene into the developing spikelets of Adh1 null maize plants. Although the approach was a canonical "long shot", the ease and rapidity with which the experiment could be performed and the results assayed by pollen staining, and the potential value of a pollen transformation system, overcame our unease regarding the rather foggy biological underpinnings of our approach. Much to our surprise, the first experiments performed suggested transformation of Adh1 null pollen to Adh +. This original result led to intermittent experiments between 1982 and 1985, first at the International Plant Research Institute in San Carlos, California and later at Eli Lilly and Company in Greenfield, Indiana, designed to test this approach.
A number of preliminary experiments were performed to investigate the optimal mechanics of the macroinjection procedure. Seed from a homozygous Adh1 null maize line (kindly provided by Dr. Drew Schwartz, Indiana University) were individually planted in ten gallon pots in the greenhouse and grown under high intensity lighting. Generally about 35 to 45 days after planting, depending on the growth conditions, a window was cut into the maize stalk just above the base of the developing tassel. The immature tassel was then gently pulled through the window and supported on a solid base for subsequent manipulations. A few young spikelets at various points along the tassel were then removed and microscopically examined for their stage in pollen development. We performed the majority of our macroinjections at a stage where the gametocytes of the central (and, therefore, furthest advanced) spikelets were in pachytene. Hence, all other spikelets injected would contain gametocytes that were at or just prior to pachytene and would, perhaps, be more likely to meiotically recombine any injected DNA into recipient chromosomes. We also found that handling and macroinjection of spikelets significantly younger than this led to death of the pollen-producing cells. We injected approximately 0.5 microliters of a solution of 0.1M glucose, 50mM Tris-HCl, 5mM MgCl2 containing 0.25 micrograms/microliter DNA. The injections were into the approximate center of individual spikelets, thus bathing the developing anthers. A hand-drawn needle with an inner diameter of approximately 15 microns and an outer diameter of approximately 150 microns was found to be optimal for the injection process. Larger outer diameters tended to kill the immature spikelet and smaller inner diameters did not allow efficient delivery of the DNA solution. All of the reasonably developed spikelets on a tassel were individually injected, any spikelets not injected were excised or marked with a felt tipped pen as uninjected controls. The young tassel was then gently replaced within the central plant stalk and the previously excised window taped back into place. Within 15 to 19 days, depending on growth conditions, the somewhat distorted tassel shed pollen. Collected pollen samples were first weighed (estimating 3000 pollen/milligram) and then stained for ADH1 activity by standard procedures (Freeling, Genetics 83:701, 1976).
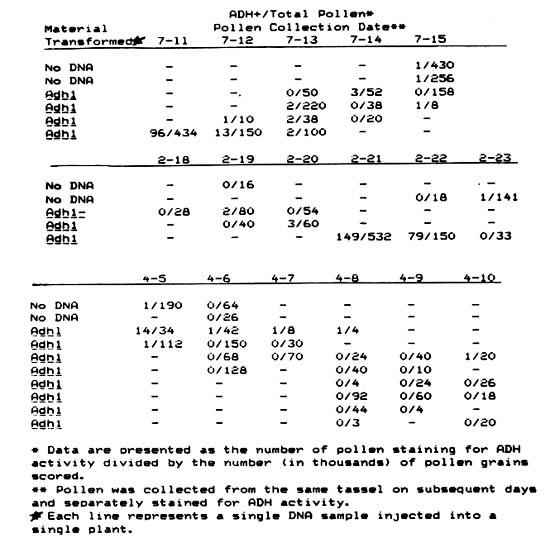
In 14 preliminary experiments, in which the individuals performing the injections and pollen scoring were unaware of the nature of the injected solutions, 1 of the 8 controls (half no DNA and half Adh - plasmid DNAs) and 4 of the 6 Adh1 injections yielded Adh + pollen. Since even a low level of contamination by pollen from an Adh1+ plant would totally invalidate our approach, we found that we could not have any mature Adh1 + plants in the same greenhouse with our injected plants and that the individual performing the pollen staining could not have recently visited a greenhouse or field that contained shedding Adh1 + plants. Also, samples of Adh1 + pollen commonly used as controls for the staining procedures could also be a major source of contamination if taken through the staining procedure by the same person or in the same room as the pollen from the injected plants. Despite all these problems, we were able to derive some provocative results when all known sources of pollen contamination were eliminated. The Table presents data derived from 3 such experiments.
In the properly controlled experiments performed, we found 19 out of 34 Adh1 injections gave an apparent transformation frequency of .005% or higher. Only 3 of 20 control injections of Adh1 - DNA or no DNA gave a similar high frequency of apparent transformants. We assume that these 3 false positives were due to mislabelling of a tube containing Adh1 + DNA as a control, from contamination by putatively transformed pollen shed in the greenhouse or from some unknown source. In no case did the controls yield more than 4 putative Adh + pollen grains in a single staining. The highest frequency of putative transformants with an Adh1 injection observed in any experiments was 0.1%.
Since we made no extensive attempt to recover putatively transformed pollen through a fertilization event, we have no absolute evidence to indicate that recovery of ADH1 + pollen from macroinjected Adh1 null plants was not due to some unidentified source of pollen contamination. The tendency of a putatively transformed plant to continue to produce ADH + pollen over 2 or more days, while those around it remained ADH -, supports a biological interpretation. Moreover, many of the putatively transformed pollen stained pink rather than fully purple, indicating a low content of ADH enzymatic activity. All Adh1 + pollen controls and contaminants consistently stained fully purple. A low level of ADHI enzymatic activity in a pollen grain could result from a transient, non-integrating transformation by Adh1 DNA or from an inefficient expression of transformed Adh1 DNA due to a structural alteration in or position effect on the integrated gene.
We eventually terminated these experiments for a number of reasons. Variations in the time of injection, the injection buffer, and the presence of various replicative or integrative components in the transformation vector did not seem to improve the transformation efficiency. In addition, we did not become significantly more successful in achieving high apparent transformation frequencies (>1/2000) with practice. More importantly, the requirement to delegate an entire greenhouse and various personnel exclusively to this project became prohibitively expensive. Had we been able to perfect a technique for reproducibly storing pollen in a viable form, we would have been able to split the pollen collected into stored and stained aliquots. Any injected plants yielding a high level of ADH + pollen could then be used in a pollination whenever the appropriate silks were available. The chief reason for the cessation of these experiments, however, was our decision to await the construction of recombinant vectors that could drastically simplify the interpretation and recovery of putative pollen transformants. These vectors, containing plant regulatory sequences that can drive gene expression in maize pollen (for instance, the promoter from maize Adh1) linked to selectable or scoreable markers not naturally encoded by maize, are now available.
J.L. Bennetzen, C. Lin, S. McCormick and B.J. Staskawicz
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}