The presence of 2 low molecular weight plasmid-like double-stranded DNA molecules, S1 and S2, in undigested mitochondrial DNA (mtDNA) preparations is characteristic of male-sterile maize cytoplasm of the S-type (cms-S). In some fertile Latin American races of maize carrying RU cytoplasm, plasmid-like mtDNA molecules, designated R1 and R2, are present. R2 is nearly indistinguishable from S2; both are about 5.4kb in length and differ in a single Bgl1 site that is missing from S2. R1, at 7.4kb in length, is approximately 1kb longer than S1 and contains about 2.6kb that are unique to R1 (R.R. Sederoff and C.S. Levings III, Gene Flux in Plants, pp. 91-109, 1985).
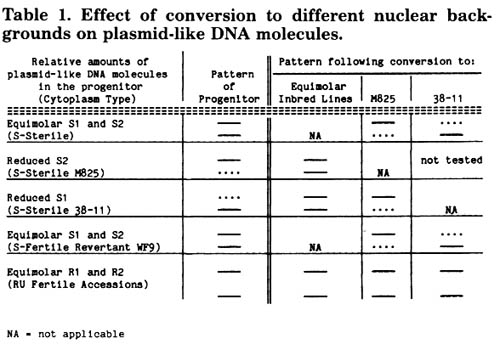
The relative amounts of the S1 and S2 episomes in cms-S strains are determined by the nuclear background. In most inbred lines carrying cms-S, S1 and S2 occur in equimolar amounts. Several inbred line backgrounds in which this is not the case have been identified. M825 and the related R825 exhibit a reduced amount of S2 compared with S1; in 38-11 the reverse is true. It was previously demonstrated that when an inbred line whose mtDNA exhibits equimolar amounts of S1 and S2 is recurrently crossed with M825 as the male parent there is a decrease in the amount of S2 as compared with S1 (J.R. Laughnan et al., Stadler Symp. 13:93, 1981). The result is the same when the inbred line 38-11 is recurrently crossed with M825; the "reduced S1" pattern shifts to the "reduced S2" pattern characteristic of M825. Not only are S1 and S2 under nuclear control, they respond to the nuclear background independently of one another. It also should be noted that while cytoplasmic reversion to fertility of cms-S in the nuclear backgrounds M825, 38-11, H95 and W182BN results in the loss of the S1 and S2 episomes, revertants in the WF9 inbred background retain S1 and S2 in the same relative amounts as do their sterile progenitor strains. Therefore a number of fertile and sterile episome-containing strains are available for studies of the effects of changes in the nuclear background.
Agarose gel electrophoresis studies of ethidium bromide-stained undigested mtDNA from various episome-containing strains converted to different nuclear backgrounds are now essentially complete (Table 1). Each strain shown in the table was crossed a minimum of 6 times to the inbred line background indicated. It has been demonstrated that 5 crosses are sufficient to shift the 38-11 pattern to the pattern characteristic of M825.
The cms-S versions of a number of strains with equimolar amounts of S1 and S2 were recurrently backcrossed to both M825 and 38-11. Following conversion to the M825 nuclear background the "equimolar" pattern of S1 and S2 shifts to the "reduced S2" pattern and following conversion to 38-11 the pattern shifts to the "reduced S1" pattern. The "reduced S2" pattern characteristic of M825 is reversible since conversion to an "equimolar" nuclear background results in equimolar amounts of S1 and S2. The backcrossing of M825 to 38-11 has been carried out but the mtDNA has not been analyzed. We predict that it would exhibit the "reduced S1" pattern based on the other results presented herein. The "reduced S1" pattern of cms-S 38-11 strains becomes equimolar following conversion to "equimolar" strains; following conversion to M825 the pattern is that of "reduced S2".
The S1 and S2 episomes present in cms-S WF9 cytoplasmic revertants retain their ability to respond to the nuclear background. Revertants converted to M825 exhibit the "reduced S2" pattern while the same revertants converted to 38-11 exhibit the "reduced S1" pattern. Whatever mtDNA changes occurred upon reversion to fertility, the S1 and S2 copy numbers are still under nuclear control. Reversion to fertility in WF9 does not alter the responses of S1 and S2 to the nuclear background.
Several RU cytoplasm accessions containing the R1 and R2 episomes were crossed by "equimolar" strains, by M825 and by 38-11. As expected, no effect of conversion to the "equimolar" strains was observed. However, conversion to 38-11 also exhibited no change in the relative amounts of R1 and R2. Since S1 responds to the 38-11 nucleus and R1 does not, this could mean that the target site for action of the 38-11 nuclear gene(s) in the S1 episome is in that region of S1 that contains no homology to R1. However, this region is present in S2 (and R2) and since S1 and S2 respond differentially to the nuclear background this is not a satisfactory explanation. This implicates a site or sites in the main mitochondrial genome as the target for the nuclear gene control. This conclusion is supported by the fact that R2 does not respond to the M825 nucleus as does S2; there is no change in the relative amount of R2 in the M825-converted RU cytoplasm. The 2 episomes, R2 and S2, are almost identical and if the target for nuclear control were present in S2 it would be expected to be present in R2 as well; results indicate that this is not the case. Restriction enzyme profiles show that the main mitochondrial genomes of cms-S and RU are very different, RU being similar to normal cytoplasms (A.K. Weissinger et al., PNAS 79:1-5, 1982). These results support the hypothesis that the target(s) for nuclear gene control of the maize episomes S1 and S2 resides in the main mitochondrial genome, not in the episomes themselves.
Table 1. Effect of conversion to different nuclear backgrounds on plasmid-like DNA molecules.
Heidi Feiler, Carol Leja, Susan Gabay-Laughnan and J.R. Laughnan
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}