Anthocyanin accumulation in maize and sorghum has been exploited in several studies as a sensitive and convenient measure of a light-controlled biological process. Physiological studies (reviewed by A.L. Mancinelli in Bot. Rev. 51:107, 1985) show that anthocyanin accumulation in these plants is an example of a high irradiance response (HIR). HIR's are characterized by:(1) continuous irradiation required to produce the response; (2) strong fluence-rate dependency; (3) response does not show reciprocity.
In maize particular allelic combinations of genes exhibit a characteristic pattern of light dependence for anthocyanin accumulation. Some of the most familiar examples include the interaction between B and pl, which gives rise to the "sun-red" effect described by Emerson in 1921 (Cornell Univ. Agri. Exp. Stn. Mem. 39). Chen and Coe (Biochem. Genet. 13:333, 1977) have described an allele (c-p) of the C1 gene which exhibits light-dependent anthocyanin accumulation in the aleurone during germination. A more recently described example is the deep purple pigmentation in the pericarp of kernels with a genetic constitution of Sn plus Pl (G.A. Gavazzi et al., Maydica 30:309, 1985). In order to produce anthocyanin in the pericarp of plants of genotype Sn plus pl, the developing kernels must be exposed to light. The light requirement for these phenomena is usually demonstrated by cutting back the overlying tissue and exposing the underlying "dark" tissue to direct sunlight or its equivalent. In approximately 24 hours the previously green (colorless, if aleurone or pericarp) tissue shows a dramatic increase in anthocyanin pigments.
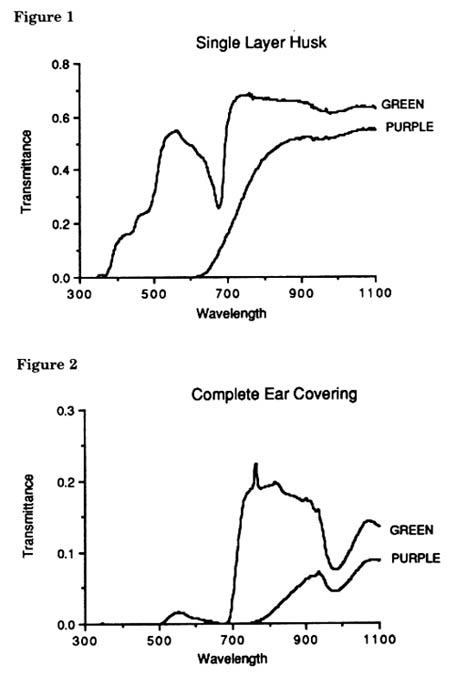
As a first step to determining the precise light requirements necessary to elicit anthocyanin accumulation we measured the light characteristics that the plants are exposed to during much of their growing season. The spectrum of sunlight transmitted through either a single layer (Figure 1) of green or purple husk tissue or through the entire intact cob covering (Figure 2) was measured with a LI-COR 1800 portable spectroradiometer in 2nm increments. Measurements were made in August and September at Stanford, CA between 1200 and 1500 hours on bright cloudless days over the wavelength range from 300 to 100nm. Transmittance was calculated as the ratio of the spectral energy distribution of each tissue to that of the incident sunlight.
A single green husk or leaf sheath (Figure 1) transmits about 20% of the incident sunlight in the blue range (420-480nm), -35% in the red range (@660nm) and - 70% in the far-red range (@730nm). In a B pl plant this is the light "seen" by the next underlying layer, i.e. the light transmitted by the newly exposed green husk. In approximately 24-48 hours, this outer husk turns purple and then transmits the purple husk pattern. Anthocyanin pigments in the husk drastically alter the amount and spectral quality of the transmitted light. Virtually no blue light is transmitted and the R:FR ratio, is 0.21 vs. 0.5 transmitted by a single green husk.
Developing kernels are exposed to virtually no physiologically relevant light below 700nm (Figure 2). An intact green ear covering does transmit light in the far-red range at levels about 1/4 that through a single layer of green husk. The R:FR ratio in an intact green ear covering is 0.14 compared to 0.005 for the complete purple covering. We have found that a single layer of green husk is sufficient to prevent pigmentation of the pericarp in Sn pl plants. This would indicate that the blue portion of sunlight must be the effector wavelength.
Loverine P. Taylor
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}