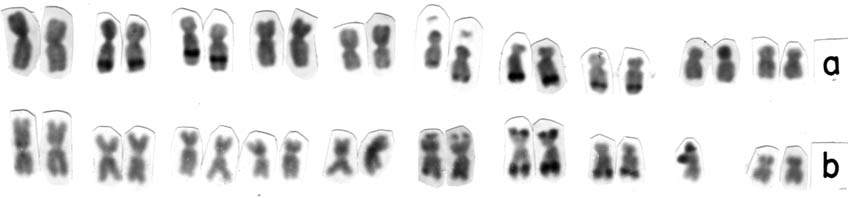
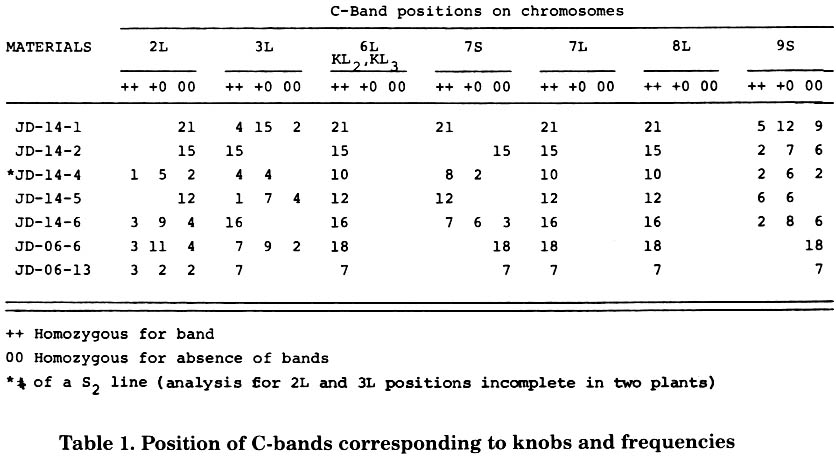
During the development of work with the purpose of selecting families of isogenic lines with slight differences in knob composition within families, a C-banding procedure was employed (Aguiar-Perecin and Vosa, Heredity 54:37, 1985). The experience has proved useful for the analysis of a large number of plants in a population with knob positions well defined. Despite the data being incomplete, it is interesting to present them. Table 1 shows the knob constitution of 2 groups of S3 progenies, JD-14 and JD-06, each one derived, from one plant of the commercial flint variety Jac-Duro (Sementes Agroceres). Data on each progeny were obtained through the examination of C-banded metaphases in root tips of seedlings from individual ears. All the materials showed homozygous bands in the long arm of chromosomes 6, 7 and 8 (Fig. 1 a, b). The analysis of pachytene showed that the band in chromosome 6 corresponds to KL2 and KL3. It has not been determined yet if the band on chromosome 8 corresponds to KL1 or KL2. So, we are using the provisional designation of "8L' for this position. Bands corresponding to K2L and K3L were also found in the two groups of lines, segregating in most of them (Fig. 1a). Only JD-14 lines displayed knobs at 7S and 9S (Fig. 1b). It can also be observed in Table 1 that the materials with more than 15 plants analyzed show the 1:2:1 expected Mendelian ratio. Deviations seem to be due to sample problems, at the present stage of the investigation. On the other hand, it is interesting that a preliminary survey of S4 and S5 ears shows that segregation is still occurring in at least one locus in most materials investigated. This is particularly consistent for S4 ears derived from JD-14-4, which, as can be seen in Table 1, have knobs segregating at 2L, 3L, 7S and 9S. Among 10 ears analyzed segregation was found in at least 2 knob positions. More progenies are under analysis and aspects like independent segregation of knobbed chromosomes will be checked in selected crosses and their F2 generations. But the possibility that in fact we have an excess of heterozygotes that we succeeded in selfing should not be discarded. This is an interesting hypothesis to be investigated, for our system could be simulating mechanisms of artificial as well as natural selection, when selection pressure for heterozygosity may occur (references in Jain, Ann. Rev. Ecol. Syst. 7:469, 1976). On the other hand, the inbreds JD-14-1 with segregation only at 3L and 9S, and JD-14-2, only at 9S, have already produced completely homozygous S6 progenies (see Table 1 of the communication on flowering time).
The lines obtained are being used in some experiments to study knob effects on development, involving flowering time screening and regeneration capacity in tissue culture experiments.
Figure 1. Karyotypes of a) plant of the progeny JD-06-13, homozygous for bands at 2L, 3L, 6L (KL2, KL3), 7L and 8L (correspondence with KL1 or KL2 not determined yet); b) plant of the progeny JD-14-1 homozygous for bands at 6L, 7S, 7L, 8L and 9S.
Table 1. Position of C-bands corresponding to knobs and frequencies of homozygotes and heterozygotes for each position in inbred lines (S3) derived from a flint maize variety.
Margarida L.R. de Aguiar-Perecin and Juliana U. Decico
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}