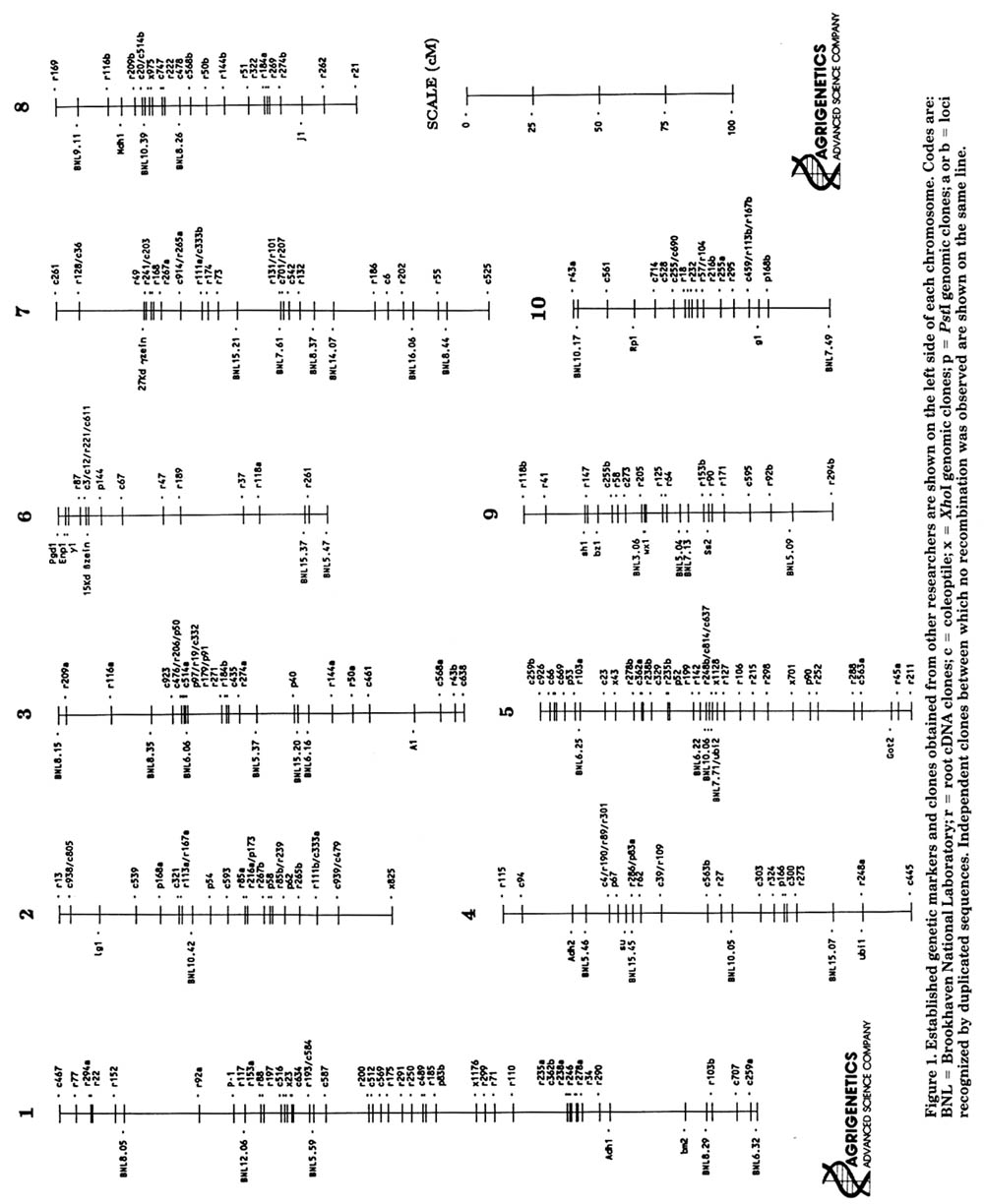
The Agrigenetics RFLP map is derived from linkage data observed in an A619Ht x Mangelsdorf's tester F2 population consisting of 87 individuals. The Mangelsdorf's tester stock was obtained from the University of Minnesota and A619Ht from Jacques Seed Co. (Prescott, WI). DNA was extracted from greenhouse-grown F2 plants which were scored for the following Mangelsdorf's tester markers: bm2, lg1, su1, y1, j1, wx1, and g1. This tester stock did not contain a gl1 marker on chromosome 7 (D. Weber, pers. commun.) and the a1 and pr1 markers were not useful in this cross. We also used three isoenzyme markers (Pgd2, Enp1, and Mdh1), several cloned genes for well-known genetic markers (Table 1) and 37 of the Brookhaven National Laboratory markers (courtesy of B.Burr). In the early stages of map development, monosomic stocks (for all chromosomes except 1 and 5) obtained from David Weber were used to confirm the location of some linkage groups.
No differences in marker linear order were detected among 75 probes used to analyze a separate F2 population (B68Ht X B73Htrhm). The r92b, r111b, r118b, r167b, c333b, Got2 and Rp1 loci were mapped in other populations. Rp1 was mapped following field inoculation and scoring of 45 F2 individuals that were segregating for Rp1.
Genetic linkage was detected using the method of expected phenotypic categories as described by Mather (The Measurement of Linkage in Heredity, Methuen and Co., London, 1938). Orthogonal coefficients for the 9-cell classification were used to determine lack of independent segregation, to examine deviation from Mendelian expectation for each marker, and to test the possibility of genetic linkage. This method reduces the number of inaccurate or physically nonexistent "mathematical" linkages which result from causes other than linkage. The method of maximum likelihood (Allard, Hilgardia 24:235, 1956) was used to calculate recombination frequencies. We have also used a method described by Burr et al. (Theor. Appl. Genet., in press) in which linear order is determined by seeking to minimize crossovers among all pairwise probe comparisons.
Table 1. Identified clones obtained from other researchers.
Identified clones obtained from other researchers#
| Cloned gene | Plasmid | Source |
| P1 | P-VB.4 | T. Peterson |
| Adh1 | pZML793 | E. Dennis |
| A1 | pALC2-2 | S. Schivarz-Sommer |
| Adh2 | pZML841 | E. Dennis |
| Sh1 | p17.6 | L.C. Hannah |
| Bz1 | pD3MS9 | 0. Nelson |
| Wx1 | pSaIC | S. Wessler |
| Ss2 | p21.2 | L.C. Hannah |
| ubi | H76 | P. Quail, A. Christensen |
| b zein | PZG15RX | B. Larkins |
| g zein | gzM5 | A. Esen |
Most of our DNA markers are cDNA clones made from two different RNA populations. The first clones, prepared from etiolated coleoptile total poly(A) mRNA are indicated by the prefix "c". These clones were constructed by G-C homopolymer tailing into the PstI site of the vector pSP64 (Promega). A high percentage (about 20%) of these cDNA clones proved unusable due to high background hybridization. Subsequent analysis suggested that excessive G-C tracts were a major cause of the non-specific hybridization. This conclusion was strongly supported when a second seedling root polysomal poly(A) cDNA library (prefix "r") was prepared by A-T tailing and less than 1% of the clones were rejected for reasons of high background. Overall, 38% of the cDNA clones were rejected for reasons of weak signals, high background or complexity. More than 80% of the useful clones were polymorphic with EcoRI or EcoR5 digests, making the use of additional enzymes unnecessary. Single locus clones were preferentially selected, since they facilitate practical utilization of the map. However, our map does include a number of two-locus markers (Table 2) which are designated "a" and "b" on the map (Figure 1).
Table 2. Chromosome locations of duplicate molecular markers.
| Chromosomes | Molecular markers |
| 1,4 | p83 |
| 1,5 | c259 c362 rl03 r235 r238 r278 |
| 1,9 | r92 rl53 r294 |
| 2,2 | r85 |
| 2,7 | c265 r267 c333 rill |
| 2,10 | rl 13 p168 rl67 |
| 3,8 | C514 C568 r50 rJ16 H44 rl84 r274 r209 |
| 3,10 | r43 |
| 4,5 | c563 ubi r248 |
| 6,9 | HIS |
| 9,10 | c255 |
Our map includes some clones derived by exploiting the apparent undermethylation of single copy sequences (Burr et al., Theor. Appl. Genet., in press). An initial attempt using 1-2 Kbp XhoI fragments cloned into a vector SalI site (prefix "x") gave only a 10-fold enrichment for single copy sequences. Subsequent libraries using 0.5-3 Kbp PstI fragments (prefix "p") were much more efficient and about 70% of these clones were mappable.
Several new genes of known identity have also been positioned on the map (Table 1, Figure 1). A 15 Kd b zein gene obtained from Brian Larkins has been mapped to chromosome 6. A 27 Kd g zein obtained from Asim Esen has been mapped to chromosome 7. A ubiquitin sequence (H76) obtained from Al Christensen and Peter Quail hybridizes to two loci (ubi1 and ubi9 on chromosomes 4 and 5 respectively). Clone r248 also proved to be a ubiquitin sequence by cross-hybridization.
Figure 1. Established genetic markers and clones obtained from other researchers are shown on the left side of each chromosome. Codes are: BNL = Brookhaven National Laboratory; r = root cDNA clones; c --- coleoptile; x = XhoI genomic clones; p - PstI genomic clones; a or b = loci recognized by duplicated sequences. Independent clones between which no recombination was observed are shown on the same line.
M. Murray, J. Cramer, Y. Ma, D. West, J. Romero-Severson, J. Pitas, S. DeMars, L. Vilbrandt, J. Kirshman, R. McLeester, J. Schilz and J. Lotzer
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}