Some mutants which condition defective kernels in maize, as we have previously studied (see MNL 56:108, 1982; 57:71, 1983 and 58:120, 1984), produce multiple morphological, physiological and biochemical modifications in seeds. Especially, the study of biochemical changes conditioned or associated with defective mutations gives opportunity to understand the nature of the mutation and the possible changes promoted in most of cases, in which the character is lethal.
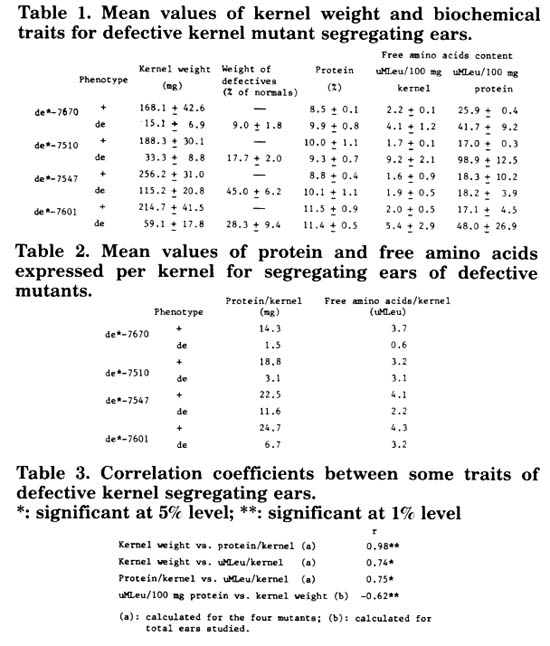
With the purpose of studying if the degree of defectiveness is associated with other biochemical characters, we have enlarged our investigations to another 3 spontaneous mutants which condition defective kernels. Four spontaneous mutants (de*-7670; de*-7510; de*-7547 de*-7601) produced in different inbred lines and conditioned by recessive genes of single inheritance were used in this analysis. Over a total of 58 segregant ears for each one of the mutations, the weight of defective and normal kernels was evaluated, determining the "defectiveness degree" through the calculation of the weight proportion of defectives relative to their normal equivalent (see Table 1).
Defective and normal kernel samples were taken at random from each ear. For each sample protein and free water-soluble amino acid content were determined. Free amino acids were expressed in equivalent leucine micro mols (µMLeu) per each 100mg of grain or protein (see Table 1).
Protein content did not differ significantly between normal and defective kernels. In contrast, highly significant differences are detected for free amino acid content in 3 of the mutants used (de*-7601; de*-7670; de*-7510), which notably affect endosperm and embryo.
Mutant de*-7547 does not affect the viability of the embryo or condition significant differences in free amino acid content.
In Table 2, results were expressed on the basis of the individual kernel. As weight decreases it can be noted that defective kernels generally have a smaller protein and free amino acid content than their normal equivalent. In the case of de*-7601 and de*-7510, this does not occur in free amino acid content. As a consequence of the high level of free amino acid content despite weight reduction, these defective kernels have almost the same free amino acid content per grain as their normal equivalent.
In Table 3 some of the possible associations that can be studied among the evaluated traits are shown. From these results, it can be deduced that the biggest kernels have a greater whole protein and free amino acid content, and relative level of free amino acid content is inversely associated with kernel weight. This fact points out that the most defective kernels have a greater percentage of free amino acids.
Three of the mutants (de*-7601; de*-7670; de*-7510) produce a strong weight reduction in kernels: defective kernels have from 1/10 up to 1/4 of the weight of their normal equivalents. These mutants are lethal as defective kernels are not capable of germinating under normal conditions, but if they do, they generate plants which die in an early stage of their development.
Defective de*-7547 does not condition such a strong weight reduction in kernels (near 50% of the weight of their normal equivalents), nor the normal viability of kernels, producing plants which come up to a reproductive stage.
The lethal mutants studied condition from 2 to 6 times more free amino acid content in defective kernels relative to the normal ones, while the viable defective does not show any difference.
The results we show in this paper point out that at least for this little sample of mutant genes that condition defective kernels the "defectiveness degree" is significantly related to free amino acid content. This phenomenon can be attributed to the next: mutant genes, we believe, produce an early blockage in the synthesis of storage proteins or a repression of determined kinds of protein, which determine that part of those amino acids translocated to kernel do not form any part of the protein fraction. Some of these non-synthesized proteins could be essential for normal embryo development. This fact would possibly explain the lethality of some mutant genes.
Table 1. Mean values of kernel weight and biochemical traits for defective kernel mutant segregating ears.
Table 2. Mean values of protein and free amino acids expressed per kernel for segregating ears of defective mutants.
Table 3. Correlation coefficients between some traits of defective kernel segregating ears. *: significant at 5% level; ##: significant at 1% level
Ida G. Palacios and Jorge L. Magoja
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}