Various cytogenetic techniques have been employed for producing whole chromosome or partial chromosome duplications in maize. However, most of the methods are difficult to utilize, since the duplications produced are not heritably stable. For example, nondisjunction of BA chromosomes can be used to construct hyperploid plants (A AB BA BA ). These plants are trisomic for a distal A chromosome segment. The condition, however, is not uniformly transmitted in crosses due (in part) to BA nondisjunction.
A procedure for constructing stable or nearly stable duplications has been developed. This method combines the chromosomes of a standard (A-A) translocation and a B-A translocation in a unique manner. The basic idea is to utilize certain adjacent I gametes produced by A-A translocation heterozygotes. Adjacent I gametes are both duplicate and deficient. They are normally inviable due to their deficiency. However, the addition of an appropriate BA chromosome will cover the deficiency and rescue the gamete along with its duplication.
The technique was tested by combining T8-9(4453) with the B9 from TB-9Sb. Translocation breakpoints for T8-9(4453) are 8L 0.86 and 9S 0.68. The adjacent I gamete to be rescued is 8 98 . This gamete lacks the distal 0.32 of 9S. In TB-9Sb, the breakpoint on 9S is 0.4 (Roman and Ullstrup, 1951). The B9, therefore, carries the distal 0.6 of 9S and can rescue the 8 98 gametic class (it also produces a second duplicated region in the gamete due to overlap between the B9 and 98).
The steps involved in producing the duplication gamete (8, 98 , B9) are as follows:
1. Cross hyperploid TB-9Sb as female to homozygous T8-9.
9(wx) 9B(Wx) B9B9 X 98(Wx) 98(Wx) 89 89
2. Germinate progeny and select Wx wx plants by pollen classification.
These plants contain 8, 89, 9(wx) 98(Wx) B9. They are heterozygous
for T8-9 and contain a B9 chromosome (the 9B was selected against in this
step. Its absence can be confirmed by a test of nondisjunction, since the
9B is required for B9 nondisjunction).
3. Cross the selected plants as female to a wx wx tester. Some
of the progeny should receive 8, 98 (Wx) B9 from the female.
4. Germinate progeny from (3) and self pollinate. Select ears that
show a reduced (less than 75%) frequency of Wx kernels (low Wx
transmission is due to pollen competition against duplication gametes).
5. From the selected ears, germinate seeds and select Wx Wx
plants. These should be duplication homozygotes (8 8 98 98 B9 B9).
6. Self pollinate to establish a homozygous stock.
By following this protocol, two plants (6799-1 and 6801-1) were identified as putative homozygotes (step 5) and self-pollinated. The seeds produced were tested to confirm presence of the duplication. The first test involved germinating five kernels from each ear and checking chromosome numbers in the roots. Results are currently available for 6799-1. All five plants derived from ear 6799-1 contained 22 chromosomes. The extra two chromosomes were shown to be B9's, as expected.
Next, outcrosses were made of the putative homozygotes. Two crosses to tester lines were selected for analysis: bz bz wx wx X 7011-2 and bz bz wx wx x 7012-6 (Family 7011 derives from plant 6799-1 while family 7012 comes from 6801-1). These crosses gave Bz progeny as expected, although an occasional bz kernel or variegated (Bz/bz) kernel was produced. Forty-six Bz progeny from the 7011-2 cross and 43 Bz progeny from the 7012-6 cross have been checked for chromosome constitution. All 89 had 21 chromosomes, as expected for crosses of a homozygous duplication. (The bz and variegated kernels have not yet been analyzed, but their appearance in these crosses suggests that B9 instability may be present).
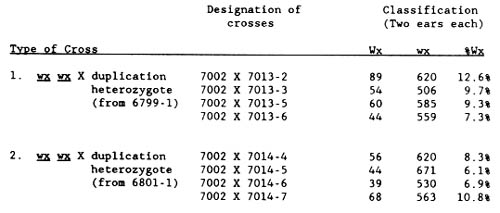
One final test was performed. Homozygotes derived from 6799-1 and from 6801-1 were crossed (as male) to a wx tester. Next, the Wx wx progeny were testcrossed as male to wx wx. Transmission of Wx through the pollen was determined. It should be less than 50%, due to pollen competition, if the duplication was present. Results for eight testcrosses are given below. In each cross, selection against Wx pollen is strong. The findings indicate that the homozygous duplication 8 8 98 98 B9 B9 has been constructed.
On theoretical grounds, the duplication is expected to be relativelv stable because a) each of the three chromosomes carries vital genes not supplied by the other chromosomes, and b) the B9 is incapable of nondisjunction in the absence of 9B. However, a potential source of instability is residual nondisjunction of the B9. The occasional (< 1%) appearance of bz kernels in crosses of bz bz x homozygous duplication indicates that some nondisjunction may occur. Also, the appearance of variegated kernels in these crosses may provide another source for changes in B9 number. However, changes in B9 dosage do not destroy the integrity of the duplication itself. A reversion to normal chromosomes can only occur by crossing over between the short, overlapping regions of 98 and B9 to restore a normal 9. The cytological results presented earlier suggest that such crossing over is not common. The n + 1 gamete is regularly transmitted by the homozygous duplication. Current tests, therefore, indicate a high level of stability of the duplication chromosome structure, even though B9 dosage may vary occasionally.
W. Carlson and R. Roseman
Return to the MNL 62 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}