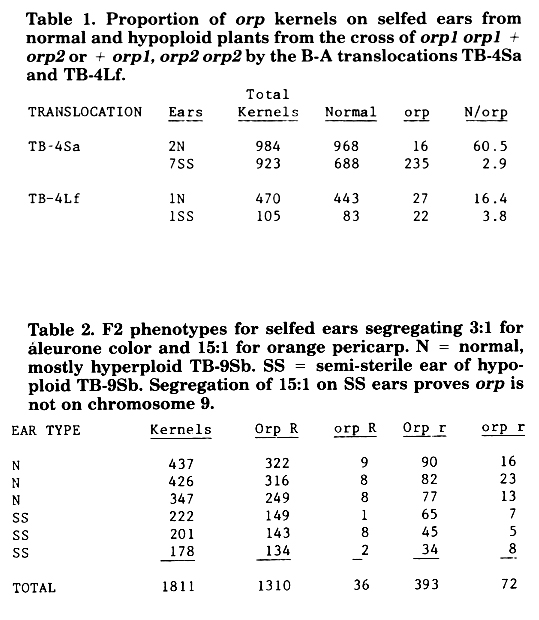
Duplicate factors conditioning orange pigment in the pericarp overlying doubly homozygous (orp1 orp1, orp2 orp2) kernels, reported last year (MNL 60:55), have now been located to chromosomes 4 and 10 respectively The first of these genes was located on chromosome 4 using a modification of the method described by Kindiger and Beckett (MNL 60:43). This was done by first crossing a stock homozygous for one factor and heterozygous for the other (from an ear showing 3:1 segregation instead of 15:1) by the full B-A chromosome set. Heterozygosity for one factor was required because the double homozygote does not survive to flowering. This material was planted and hypoploids were identified for each translocated arm. Hypoploids plus a few normal plants in each case were selfed and the resulting ears were examined for 15:1 vs. 3:1 segregation for orange pericarp. The former would occur in the double heterozygote +/orpl, +/orp2 hypo/( ) where the mutant was not on the hypoploid arm. The latter would occur when the factor was located on the hypoploid arm orp1/( ), +/orp2. Hypoploid selfs for all but TB-4Sa and TB-4Lf gave the expected 15:1 segregation (TB-10Sc and TB-10L19 were not tested). Two families of TB-4Sa gave 18 semisterile ears (hypoploids) of which 11 had an apparent 3 normal : 1 orange segregation (the other 7, as well as the nonspecified ears below, did not segregate), and 8 normal ears of which 4 had a surprising 61 normal: 1 orp segregation. One family of TB-4Lf gave 6 semisterile ears of which 1 had a 3:1 segregation, and 9 normal ears of which 4 had a 15:1 segregation (a summary of counts of some of these ears appears in Table 1). The difference in ratios on the normal ears (4S, 60:1 and 4L, 15:1) from plants that probably were hyperploid for their respective arms proves that one member of the duplicate pair is actually located on 4S and not on 4L.
Fortunate circumstances provided the location of the other member on chromosome 10 even though the B-A crosses for 10S and 10L failed to give segregating ears because a homozygous normal female was selected. In several of the selfed ears from the other B-A crosses a segregation for aleurone color due to R occurred. Classification of orange pericarp on colored kernels is difficult but possible by examining the pericarp overlying the embryo. An examination of selfed ears from normal plants segregating 3:1 for aleurone color and 15:1 for orange pericarp revealed an excess of orange kernels in the colorless (rr) class (Table 2). This indicates that the selfed plants were orp r/Orp R with r and orp linked in coupling, and provides F2 data from which an orp r map distance may be derived. The segregation ratio of R:r was 2.9:1 and for Orp:orp was 15.8:1, which is not significantly different from the expected 3:1 and 15:1. The map distance calculated by maximum likelihood is 19 ± 3 centimorgans.
In view of these findings we have designated the duplicate factor on 4S as orp1 and the one on 10 linked to r as orp2.
Table 1. Proportion of orp kernels on selfed ears from normal and hypoploid plants from the cross of orp1 orp1 + orp2 or + orp1, orp2 orp2 by the B-A translocations TB-4Sa and TB-4Lf.
Table 2. F2 phenotypes for selfed ears segregating 3:1 for aleurone color and 15:1 for orange pericarp. N = normal, mostly hyperploid TB-9Sb. SS = semi-sterile ear of hypoploid TB-9Sb. Segregation of 15:1 on SS ears proves orp is not on chromosome 9.
M.G. Neuffer, J.B. Beckett and Allen Wright
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}