There are 5-6 loci in maize known to confer resistance
to rust disease caused by P. sorghi. The Rp locus maps at the tip of the
short arm of chromosome 10 and has 14 alleles, Rp-a-Rp-m. Each allele can
be recognised by the appropriate race of the pathogen and by reaction type,
e.g.:
| Race 1 | Race 2 | |
| Rp-d | -(;)* | +(4) |
| Rp-g | -(o) | -(o) |
| Rp-m | +(4) | - (1) |
*The symbol in brackets describes the reaction using the terminology developed for wheat where ; (fleck) or O (immune) are the resistant reactions and 4 is fully susceptible.
Our original experiments were designed to tag the Rp gene with a known controlling element in order to provide a mutant accessible to molecular analysis. The Rp-g allele was the first target selected because the immune or (o) type reaction is the most clear cut resistance phenotype. In initial experiments with transmission through either pollen or egg the Rp-g allele gave rise to Rp-g' variants that were fully susceptible and occurred at about 7/1000 seedlings screened. Recovered Rp-g' alleles were stable and did not revert back to the resistant phenotype in any of 11,716 seedlings tested. Seven independently occurring Rp-g' alleles combined in all 21 possible heterozygous combinations showed no complementation. The frequency was unaltered by the presence of any known controlling element system and presumably reflects some inherent instability of or event at the Rp-g allele. The frequency was clearly several orders higher than would be expected for an insertion event and ruled out the use of this Rp-g allele as a target for gene tagging. A subsequent examination of other alleles indicated that this was not an isolated event.
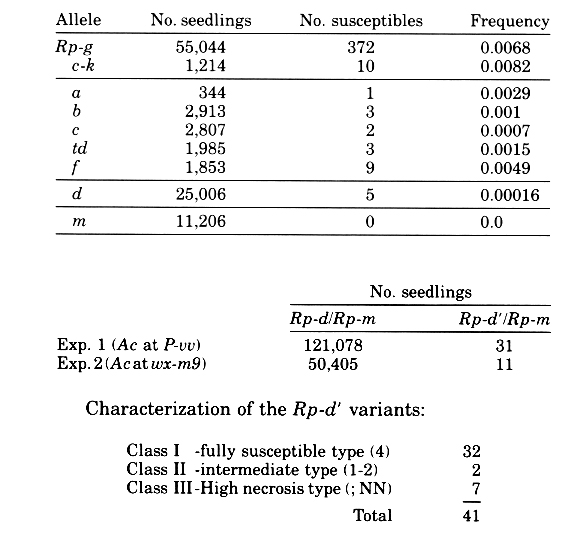
The stabilities of Rp alleles fall into four frequency classes. The Rp-m allele shows the highest stability with no clear case of an Rp-m' susceptible variant being recovered from 11,206 seedlings scored. However, the resistance reaction is a type (1) or even (2) and this is difficult to score under some conditions.
The Rp-d allele, the next stable allele, was used as a target for tagging with the Ac-Ds system. If the background stability of Rp-d is about 1-2/10,000 and the expectation for controlling element insertion is assumed to be in the range 10-4 to 10-5 then in a population of 2-300,000 seedlings one would expect to recover 40-60 Rp-d' variants due to the background instability of the Rp-d allele and 1 or 2 susceptible seedlings due to insertional inactivation by Ac-Ds. These insertional events can only be distinguished if they are unstable and revert either to resistance or some other recognizable phenotype. Cross: Rp-d/Rp-d, Ac/- x Rp-m/Rp-m
Forty of the 41 variants were recovered and are being tested for stability in the presence and absence of Ac. Classes II and III were not observed in the control (no Ac) population of 25,006 seedlings. One of the Class Il variants, Rp-d'-5, is an unstable phenotype giving rise to fully susceptible type 4 seedlings at a frequency of 0.4% (13/2725). This frequency of instability is in the range expected for a Ds excision event but further work is required to demonstrate that it is Ac dependent.
Analysis of the Class III high necrosis mutants is only preliminary. However, they appear to be very similar to barley ml-0 mutants and to the lesion mutants described by Neuffer. Rp-d'-21, a class III variant, reacts to all races of the pathogen tested and this high necrotic reaction is induced spontaneously under low temperature conditions. Two of Neuffer's Les mutants (Les*-1451,Les*-1453) are located on the short arm of chromosome 10 and may be allelic to Rp. Class I and III variants could be explained if the Rp gene contained at least two parts, one conferring specificity (S) and the other triggering the hypersensitive response (HR). Mutations in HR would be incapable of response and phenotypically susceptible, while mutants in the S part would recognise all races of pathogen (as for ml-o and Rp-d'-21) and under environmental extremes may lack control and spontaneously give rise to necrotic lesions. Such a model would explain the existence of the estimated 60-70 lesion loci in maize since they would represent the evolutionary library of resistance genes that function by initiation of the hypersensitive response.
Tony Pryor
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}