In MNL 58:38-46 the authors detected kernel row number factors Krn in all chromosomes except 4, which was not tested, and 10, which was not significant. In MNL 59:23-24 we gathered evidence of one more in 4S. In MNL 60:33-34 we pointed out that probably in 1 and 4 there was a pair of Krn factors. In the preceding work with Portugues Fasciado (PF) we found a Krn in 4L, and confirmed a few others already known. The new data also showed that fasciation in the sample tested was detected nearly as frequently as Krn. W.C. Galinat (in D.B. Walden, ed., Maize Breeding and Genetics) in the item on two-ranked versus many-ranked, reports tr in chromosomes 1, 2, 3, 6, 7, 8, 9, and 10. In the item on single versus paired female spikelets he reports pd in 3, 4, 7, 8 and 10. With the data from PF and Durango teosinte it dawned on us that we had unconsciously done with cultivated maize what had been done with wild maize, that is, teosinte. So Krn is probably allelic to tr and its symbol should turn to Tr. Fas, fasciated, is conceivably allelic to pd and would turn to Pd. With these data we arrive at 12 pairs of alleles for kernel row control in the species, linked to Fas in 4, 5 and 8 and to pd in 3, 6, 8 and 10 in total 3, 4, 5, 6, 8 and 10. See prior report with reanalysis of Langham's Durango teosinte data. Botanists have considered these types of data as incongruous. If we accept that wild maize was domesticated independently in three or four places, or if, not fully domesticated, it was transported into the presence of other races of teosinte before the isolating knobs were fully formed the story becomes congruous. Teosinte didn't transfer the knobs to maize, both developed the knobs as an isolation mechanism within each to eliminate cross pollinated individuals.
Looking at a map of the knobs in the species one is struck by two very marked characteristics. The knobs are wide apart or they are in three clusters of two to four knobs, in #6 three, L1, L2, L3, in #8 L1, L2, and in abnormal #10, four unidentified knobs.
It is as if the knobs inhibit the formation of adjacent knobs in one isolated race, and so for instance in #6, L1, L2 and L3 would be from three different domestications. B. McClintock (in D.B. Walden, ed.) calls attention to the distribution of a small knob at position three, L3 in #6, which with one in 7L she defines as the "Andean Complex". That takes care of three of the six lineages proposed by PC. Mangelsdorf in Corn, Its Origin, Evolution and Improvement. These lineages are Chullpi, Kculli, and Confite Morocho. So, for L3 in #6 the question is settled. There remain the lineages Palomero Toluquen6, ChapaloteNal Tel, and Pira Naranja and the other knobs.
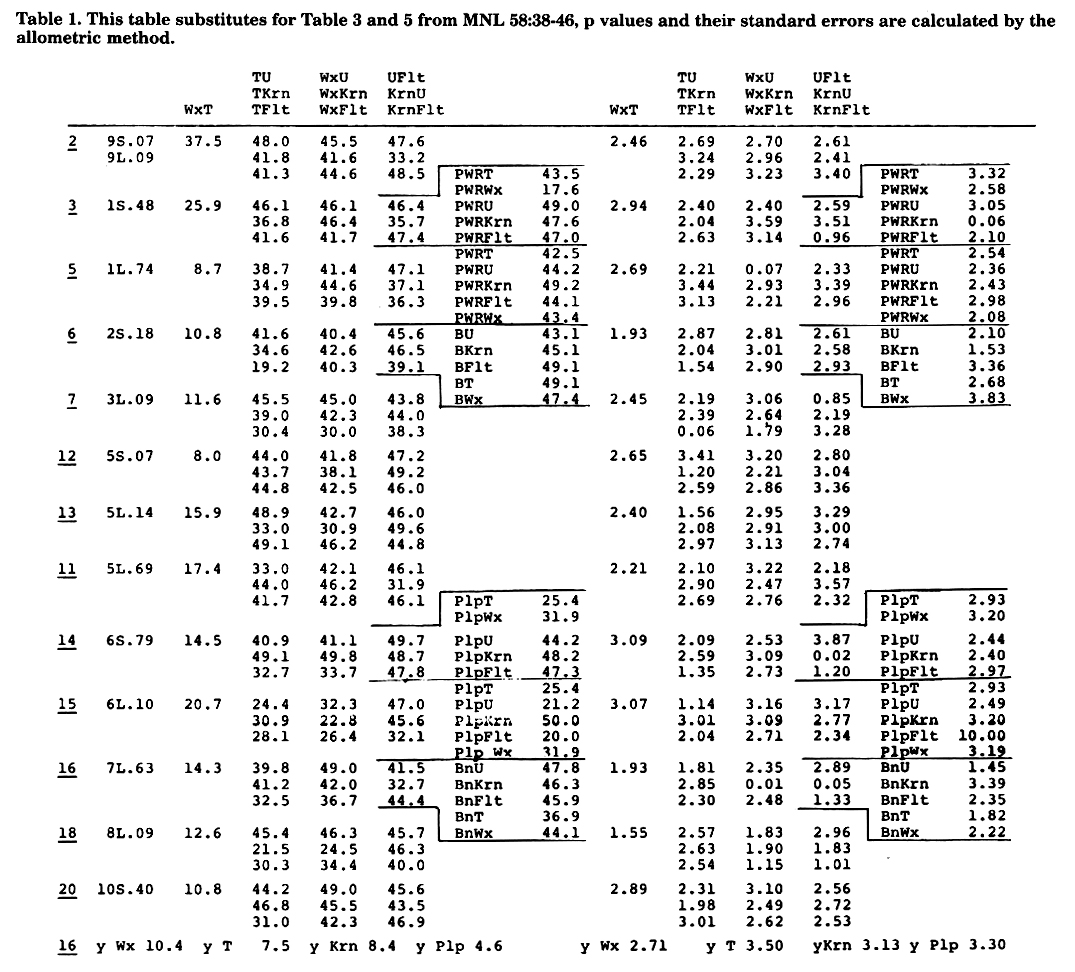
The development of the allometric method to calculate p values, shown in prior reports in this issue, led us to recalculate the data presented in MNL 58:38-46. The new results are presented in Tables 1 and 2.
Discussing the results from Table 2, we will make reference to the prior work on Durango teosinte (DT) and Portugues Fasciado (PF). The linkages detected for Krn1 in #1 seem for a locus different from those detected in DT which then should be named Tr11 and Pd11. For #2 there is agreement with DT, and with the work mapping Lsc which gave (Krn2 10) (lte1 30) (Lsc 35) (B 49). For #3 there is agreement with Tr3 from DT. For #5 we verify that for 5S.07 the sequence is Wx T and there are different loci which then would be Krn15 and Fas15, it agrees roughly with DT. For #7 there is agreement with PF. For #8, best fit would be the order Wx T for 8L.35 in PF and not as is shown in the Table.
Turning again to PF data 4L.33 could check with Ga-S Krn4 in MNL 59:23-24, and 4L.90 involves new loci, presumed Krn14 and Fas14, but with very small families. For #9 there is a reference to tr in the literature. As there is a reference for tr pd in the literature we include our sequence Flt10 Krn10 Ger5 which was not presented in the first old report.
It goes without saying that much of this summing up is quite tentative but we think it will encourage more precise work with much more powerful tools for this kind of factors.
Luiz Torres de Miranda and Luiz Eugenio Coelho de Miranda
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}