The authors (MNL 58:38-46), using 13 of the adapted standard translocations, T wx and Cateto Palha Roxa, reported positions and distances in centimorgans involving T, wx, Krn (kernel row number), Flt (flint against dent endosperm), and Ger (glucoside earworm resistance, which includes Zer and Mer). Now results are reported from crosses of 7 of the adapted standard translocations with a white dent variety with high kernel row number, Portugues Fasciado (PF, Fasciated Portuguese), probably HB 19.
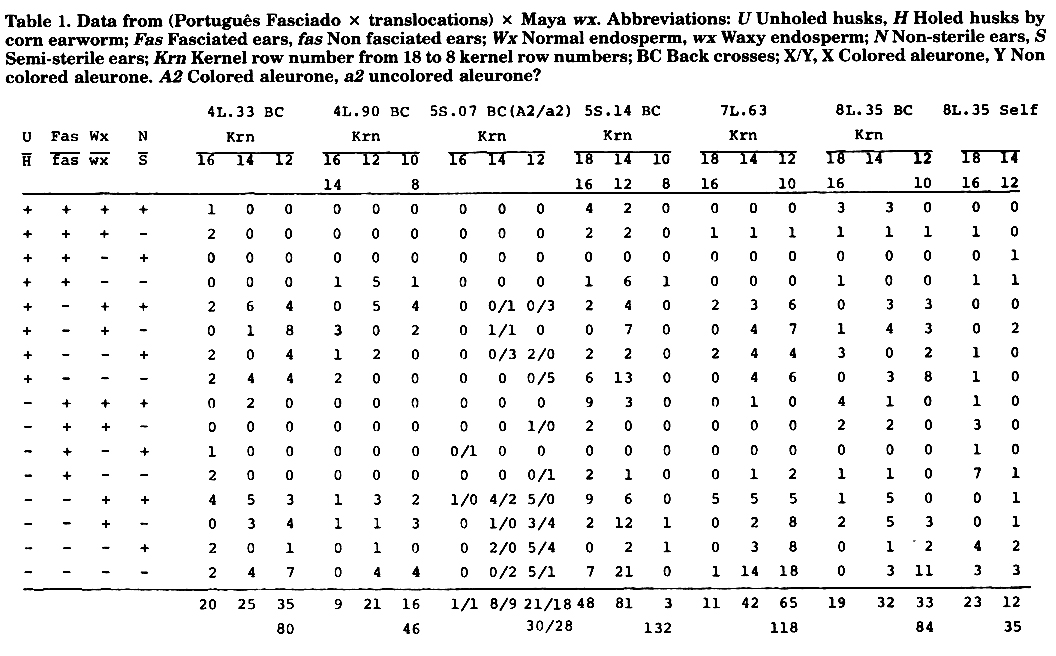
S.E. Pego in a Ph.D. thesis (Genetic potential of Portuguese maize germplasm with abnormal ear shape, Iowa State University, 1982), studied PF. He concluded fasciation is not associated with ra1, ra2 and ra3 (ramosa), and a complex system of inheritance was proposed. In our experiment in direct comparisons PF gave 16.0 mean kernel row number, while our local IAC Maya gave a 12.3 mean. The cross with translocations was backcrossed to IAC Maya wx, and in one case we also obtained results from a self. The field plots consisted of one row 10m long planted in hills 0.4m within the row, with rows 1m apart. The hills were planted with three seeds each, alternating hills with normal and waxy endosperm of the same ear within the row. Six reps were used. Every fifth row was planted with IAC Maya waxy as pollinator and the experiment was detasseled. The dry ears were harvested indiscriminately, unhusked. They were classified for presence and absence of exit holes of corn earworm, for fasciated vs. non fasciated, for Wx vs. wx, for normal vs. semi-sterile ears, and kernel row number. In the crosses with the translocation at 5S.07 there was a segregation for aleurone colour which turned out to be due probably to A2 and a2 segregation. If anybody noted this in the original translocation material please report in MNL or write to us. Although the initial stand should be up to 450, leading after thinning to 300 plants at harvest, we got analyzable data from families of only 132 to 35 plants. This was due to the very low adaptation of PF The plants simply died out. This is important to note because we used all treatments up to this number. We did not choose the results presented. At flowering time the overall conditions were unfavorable leading to very bad pollination. This explains the difficulty in separating normal from semi-sterile ears, leading to loose direct values of p for the T wx linkage. The results are presented in Table 1. In Table 2 the X2 analysis is done. The p values are presented in Table 3. The p values were calculated by the allometric method presented in a prior work, the formula being:
The p values were transformed into cM and using our "theorem of the differences" we got the cM values as the mean of the k-2 estimates available, k being the number of factors including T.
We made a modification in relation to the MNL 58:39-46 report in taking the algebraic mean of the differences, and not the mean of their absolute values as in the prior work. The distances, with their standard error, are presented in Table 4. Here we worked out the variances of the estimates.
Vp was calculated by the classical method since we saw by the exact allometric method variance formula the 2 added in the numerator is balanced by the value within brackets in the denominator which is very near 2. The covariances are a much more important component of the error estimate. To transform Vp to VcM we make VcM = (CM/p)2 X Vp. A first variance of the measure obtained by differences is the sum of the four VcM used in the two contrasts, divided by the square of the number of contrasts (two squared). As this doesn't take into account the covariances with the four cM values, we calculated a second new variance and a covariance. Remember Va+b = Va + Vb - 2 Covab. With this we know the proportions and signal of covariance, and the variance. With this proportion we correct the first variance calculated. If the covariance is positive it will be diminished, if negative increased.
In estimates of T 8L.35 backcross and selfs the p values were combined by their quantity of information. For 5S.07 the estimate Fas Krn was not utilized in the calculus. It is apparent that, as for kernel row number, factors for fasciation are as, or nearly as, ubiquitous. Really it seems they go around in pairs as tr, two ranked, and pd paired to which they are probably alielic, domesticated and wild ones.
At loose linkages allometric and classic methods gave very similar results. There is an increasing difference as p tends to zero. This is the same as reported for the F2 case.
Table 1. Data from (Portugues Fasciado x translocations) x Maya wx. Abbreviations: U Unholed husks, H Holed husks by corn earworm; Fas Fasciated ears, fas Non fasciated ears; Wx Normal endosperm, wx Waxy endosperm; N Non-sterile ears, S Semi-sterile ears; Krn Kernel row number from 18 to 8 kernel row numbers; BC Back crosses; X/Y, X Colored aleurone, Y Non colored aleurone. A2 Colored aleurone, a2 uncolored aleurone?
Table 2. X2 analysis from data of Table 1.
Table 3. p value calculated by the allometric method, with their accompanying allometric effects a and b with the data from Table 1. Extra data from 5S.07 at table bottom. Values within brackets are by the product moment method since by the allometric method the programmed calculator did not give a solution.
Table 4. Linkage sequences in cM of T wx Krn Fas in crosses of Portugues Fasciado with the indicated translocations. p values of Table 3 were calculated by the allometric method. Then it was transformed to cM and the distances arrived at by the theorem of the differences (see MNL 58:38-46), taking the algebraic mean of the differences. Variances were gotten by first transforming Vp to Vcm which is Vcm = (cM / P)2 X Vp. A first error estimate is the sum of the four Vcm divided by 4 (two contrasts squared). In 5S.07, six Vcm divided by nine (three squared). The relative size and sign of the covariances was calculated with the cM values used and subtracted two times if positive or added if negative, to the first error estimate.
Luiz Eugenio Coelho de Miranda and Luiz Torres de Miranda
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}