In MNL 58:46-48 and 48-50 we demonstrated linkages of the Latente loci in a Cateto line: Lte2 in chromosome 10 near position 56, and Lte1 19 units from B in chromosome arm 2S. This was shown by heat tolerance for the first locus and by aluminum tolerance for both loci. For heat and aluminum tolerance the Cateto system depends on the action of two pairs of complementary dominant genes. So far the latente-1 from Michoacan has shown effects only on chromosome 2 (see also MNL 56:28-30). Using the heat test lte1 is linked with wx T2-9b and Inv2a, and not with wx T9-10b. So far our results show that the Cateto Lte1 Lte2 is dominant or epistatic over the Michoacan lte1 system.
In crosses with non-Cateto flint, and dents, the lte1 is dominant. The latter statement is confirmed by the results of R.M. Castleberry and R.J. Lerette (Proc. 34th Ann. Corn and Sorghum Res. Conf., 1979). These germplasms have a bottom-most allele, for which we propose the symbol lte-0, or lte-b, for bottom.
L.T. de Miranda, L.E.C. de Miranda and E. Sawazaki in Ecological Genetics and Maize Breeding in Pt, Cargill Foundation ed. 30p. 1984, Campinas, SP, Brazil, and 0. Brunini, L.T. de Miranda and E. Sawazaki in the Conference Internationale, Paris 11-14 September 1984, Les besoins en eau des cultures, in two papers in English, p. 205-212 and 361-368, present extensive comparative measurements of physiological effects in latente-1 and other maize genotypes. We suggested lte1 has near it a factor for stomatal control, which is the object of this report.
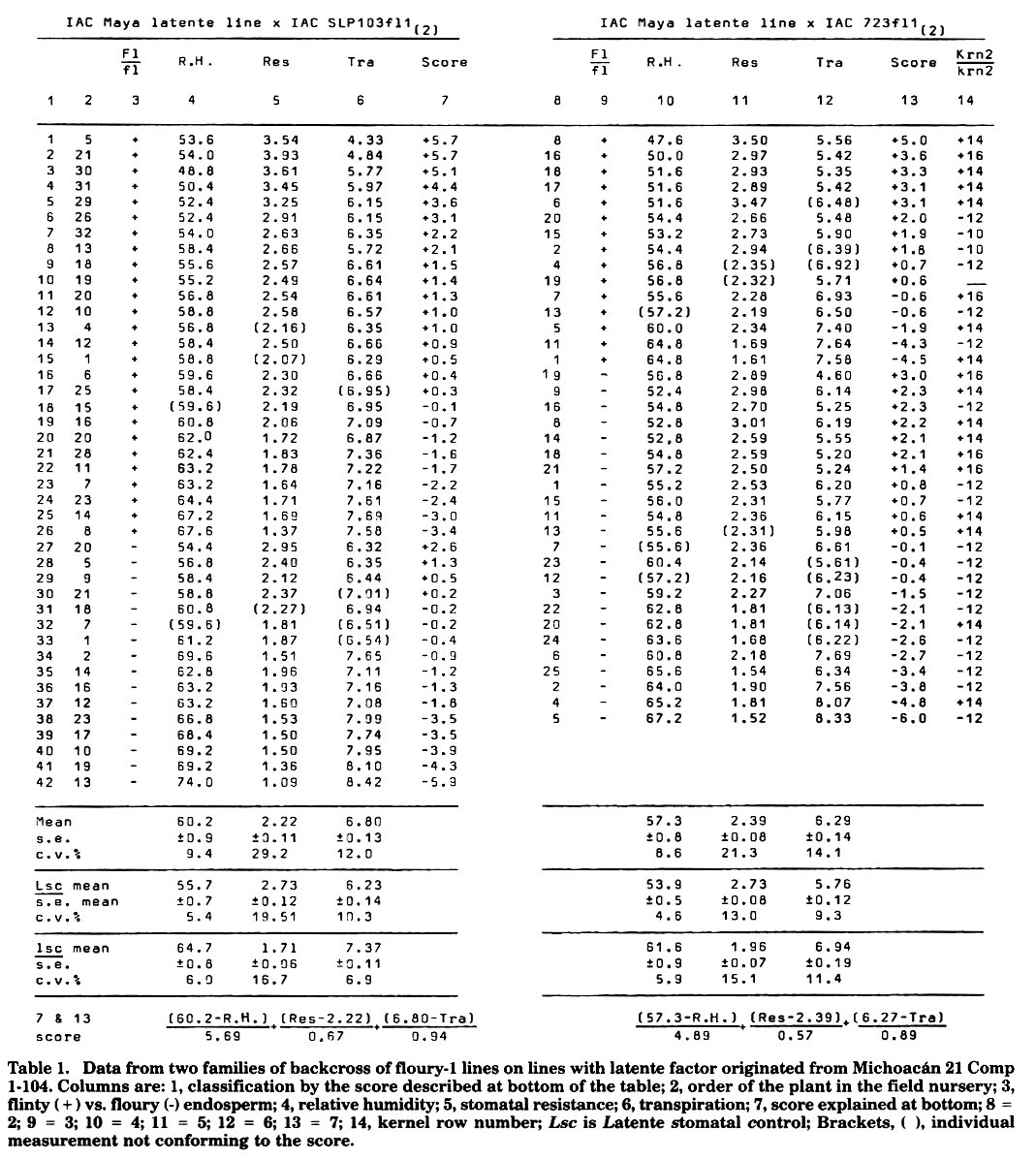
A floury-1 originally received from Pennsylvania was crossed and backcrossed four more times before selfing to the dent lines IAC SLP103 (which is an old line selfed from the Yellow Tuxpeno collection San Luis Potosi 103, from former Oficina de Estudios Especiales, now CIMMYT) and IAC 723 (an old line selfed from Tuxpan, a Tuxpeno variety from Texas, reportedly obtained by introgressing yellow colour from Creolle Yellow Flint to a typical White Tuxpeno). So we had SLP103fll(5) and a 723fll(5). These fl1 lines were crossed and backcrossed to two different Fl1 lte1 IAC Maya latente lines. The Fl1 and fl1 seeds we planted in split-plot in the field nursery in Campinas, SP, Brazil. The readings were done between 1400 and 1600 since at this period the papers cited show the greatest differences among genotypes, on March 17, for the first and March 18, 1986, for the second family. The measurements were made 10-20 days after pollination, preferably on the ear leaf or the next below or above, depending on which was healthier. The equipment used was a Licor 1600 with which we measured percent relative humidity at equilibrium of the chamber with the leaf blade, leaf stomatal resistance (s.cm-1), and transpiration in µg dm-2 s-1. The measurements were made in zig-zag, alternating whenever possible in Fl1 and fl1 rows. The results and part of the analysis are shown in Table 1. The three physiological measurements made it possible to build a score for latente contrasting with the mean to give positive values for latente characteristics and dividing each by the standard error. The highest positive values give a greater degree of ascertainment for latente and the highest negative values more ascertainment for non-latente. In the harvested ears the kernel row numbers were counted. The X2 results for 103, 723, the total and the interaction results respectively were, for Lsc1 and Fl1; 4.92*, 1.30, 5.80* and 0.42 with p = 29.6 ± 5.0, p = 40.4 ± 8.0 and mean of p = 35.0 ± 5.3. For Lsc1 and Krn2 only the second line gave indication with X2 = 2.78 and p = 29.7 ± 7.4. For Fl1 Krn the same line gave X2 = 0.30 and p 45.3 ± 8.1. The values presented were calculated by the product moment method since by additivity they give looser results. Transforming the values to map units we have the distances Lsc Krn = 34.3, Lsc Fl = 53.1 and Krn Fl = 67.2, which would lead to a Krn Lsc Fl1 sequence. We think a more correct value will be arrived at by applying the "theorem of the differences" as illustrated empirically in MNL 58:38-46 with the standard wx translocations. In the present case the distance Lsc Krn is obtained by Krn Fl1 - Lsc Fl = 67.2 - 53.1 = 14. 1. Lsc Fl by Krn Fl - Krn Lsc = 67.2 - 34.3 = 32.9. We get Krn 14.1 Lsc 32.9 Fl. Subtracting from 32.9 the distance B Fl (19.0) we get 13.9 as the distance Lsc B, which agrees nicely with the previously reported lte1 B equal to 19.0. So latente-1 must be near position 30, and Lsc in 35. Lsc and lte1 must be very near because of epistatic effects. There is no profit in closing the stomata more at highest heat, if you do not also have higher heat tolerance. The estimation of the relative position of Krn will agree with our previous work only if the directions of the genes were inverted in the first report, unless there is another gene for kernel row number, which is less probable. For B Krn distance, reported as 39, we now have 28. Note that both latente loci are very near mutagenic factors, lte1 Lsc1 near Mut, mutator, and Lte2 near cm1, chloroplast mutator. This must have an evolutionary meaning.
More precise location and study of lte1 Lsc probably will be obtained using lg1 gl2 and also leaf water potential measurements, since latente-1 has 3-4 Bars less negative leaf water potential at all times of day. If or when available, isozymes of the abscisic acid biochemical pattern should be included, as should the substitution of cysteines responsible for the bridges of sulfhydryl-disulfide bonds (SH-SS), to understand its fine structure.
Luiz Eugenio Coelho de Miranda, Luiz Torres de Miranda and Orivaldo Brunini
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}