I would like to summarize my thinking about the differentiation and functional organization of the shoot apical meristem of maize. The analysis of cell lineages during embryogenesis shows that the planes of early cell division neither specify the axis of bilateral symmetry nor demarcate the separation of major parts of the embryo or plant (R.S. Poethig et al., Dev. Biol. 117:392, 1986). This observation and the fact that more than one pattern of cell lineages during embryogenesis can be present in a plant strongly argue against the developmental significance of early cleavage patterns. In carrot, cell lineages of zygotic embryos correspond to Solanad type while those of the somatic embryos correspond to the Onagrad type (A.A. McWilliams et al., Ann. Bot. 38:243, 1974). In plants, by and large, the position of a cell rather than its lineal descent seems to determine the cell's fate.
The progenitors of the plumule-radicle axis become morphologically distinct approximately 10 to 12 days after pollination in the transition stage embryos (L.F. Randolph, J. Agr. Res. 55:881, 1936). The determination of the shoot meristem occurs just prior to or during its differentiation (Poethig et al., 1986). Auxin transport in immature embryos is known to be strictly basipetal and during embryogenesis physiologic polarity (auxin transport and auxin-cytokinin ratio) precedes morphogenetic polarity (S.C. Fry and E. Wangermann, New Phytol. 77:317, 1976; T. Przybyllok and W Nagl, Z. Pflanzenphysiol. 84:463, 1977). It is reasonable to propose that because of polar transport, auxin accumulation at the radicle end could result in the induction and determination of the root meristem. The number of embryonic cells giving rise to root meristem is not known at this stage. Auxin and cytokinin could also be involved in the determination of the shoot meristem.
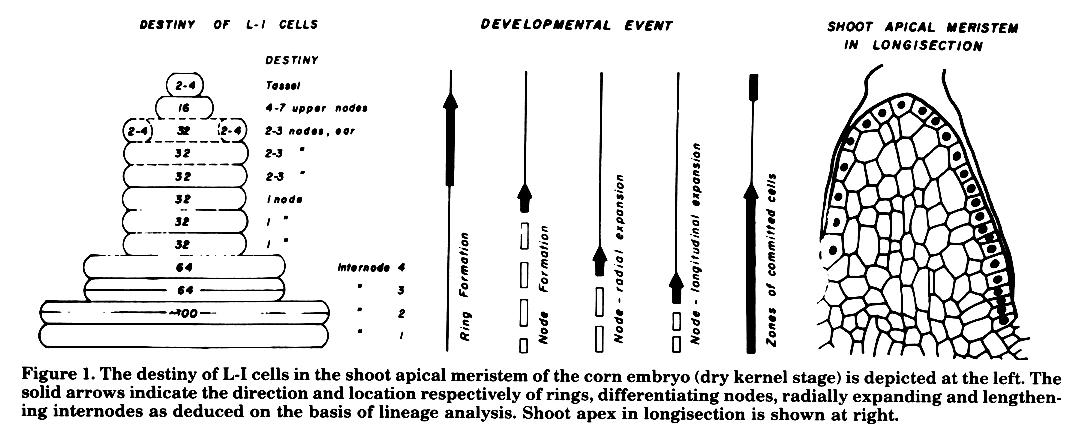
The fate of cells becomes progressively more restricted. The restriction process proceeds centripetally starting in the presumptive meristem field during the determination of the shoot meristem (Poethig et al., 1986) and continuing in the cells of the shoot apex during the formation of individual nodes (Figure 1).
The destiny of L-I cells of the apical meristem is shown schematically in Figure 1. These are two zones where cells have become committed to produce specific structures. The cells at the distal end and at the base are committed to produce, respectively, the tassel and the lower 6-7 nodes. The cells in between the two zones are relatively uncommitted and show a gradation of proliferation capacity. What does this gradation signify? Clearly these cells are in transition from an uncommitted state (i.e., cells of the ring) to become committed (the committed state being represented by cells destined to produce a specific node). The gradation therefore reflects the successive steps on the way to node differentiation from a 'ring'. While the embryo is developing, the potential of the cells committed to produce a node starts getting expressed and leads to the formation of leaves and internodes. The formation of nodes can be thought of as a wave of commitment process progressing towards the distal end. The development of the corn plant thus involves three successive and interrelated events, (a) formation of 'rings', (b) a commitment process to produce single nodes, and (c) expression of this commitment leading to the development of internodes. These events are separated from each other temporally and spatially only slightly and the shoot apical meristem represents a structure where the above events have been frozen at a specific instant of time in development.
To present an overview, it is visualized that first a rough outline of the shoot is laid down. This outline consists of tassel initials and a stack of 'rings' each of which is capable of producing a group of nodes. Then in the later part of embryonic development and continuing during the post-germination phase, a refinement process seems to occur. This process results in the production of specific individual nodes.
M.M. Johri
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}