In 1983, Robertson reported (Mol. Gen. Genet. 191:86-90, 1983) that inbreeding Mu stocks would result in the inactivation of germinal mutator activity. Others have shown that this is accompanied by modification of the Mu1 and Mu1-like elements present in these stocks (Bennetzen, Plant Genetics, M. Freeling, ed., pp. 343-353, 1985). Similar Mu1 element modification has been shown to accompany the loss of somatic mutability in Mu-induced aleurone mutants (Chandler and Walbot, PNAS 83:17671771, 1986). Thus, it seemed reasonable to assume that inbreeding a Mu-induced aleurone mutant would eventually result in the loss of mutability accompanied by modification of Mu elements.
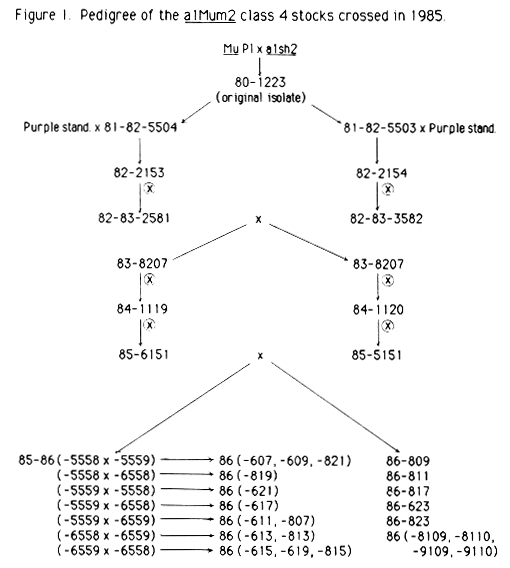
In 1985, crosses were made between two different stocks of a Mu-induced a1 mutable mutant, a1-Mum2, which exhibited a consistent expression of an intense mutable pattern (mutability scale used in this report: Class 1 = stable, Class 2 low mutability, Class 3 = medium mutability, Class 4 high mutability, Class 5 = purple). With the 1985 cross there had been 5 generations of inbreeding (either selfing or intercrossing, see Figure 1). This is equivalent to Mu32 per se in our previous inbreeding experiment (Robertson, Mol. Gen. Genet. 191:86-90, 1983). In the previous work, modification and loss of germinal Mu activity had been observed to be complete by Mu16, at least one generation earlier than that of the a1-Mum2 stocks crossed in 1985. The inbreeding in the a1-Mum2 stock has been more intense than in the previous work because it involved 3 generations of selfing which was never practiced in the original inbreeding experiment.
Sixteen ears were produced as a result of intercrossing in 1985. Seven of these had nothing but Class 4 seeds; six had predominantly Class 4 seeds with a few Class 2; one had predominantly Class 4 with a few stable seeds (Class 1) and two ears had a spectrum of mutable classes. On one of the latter two ears, the mutable classes ranged from Class 4 to Class 2. On the other, the range was from Class 4 to Class 1.
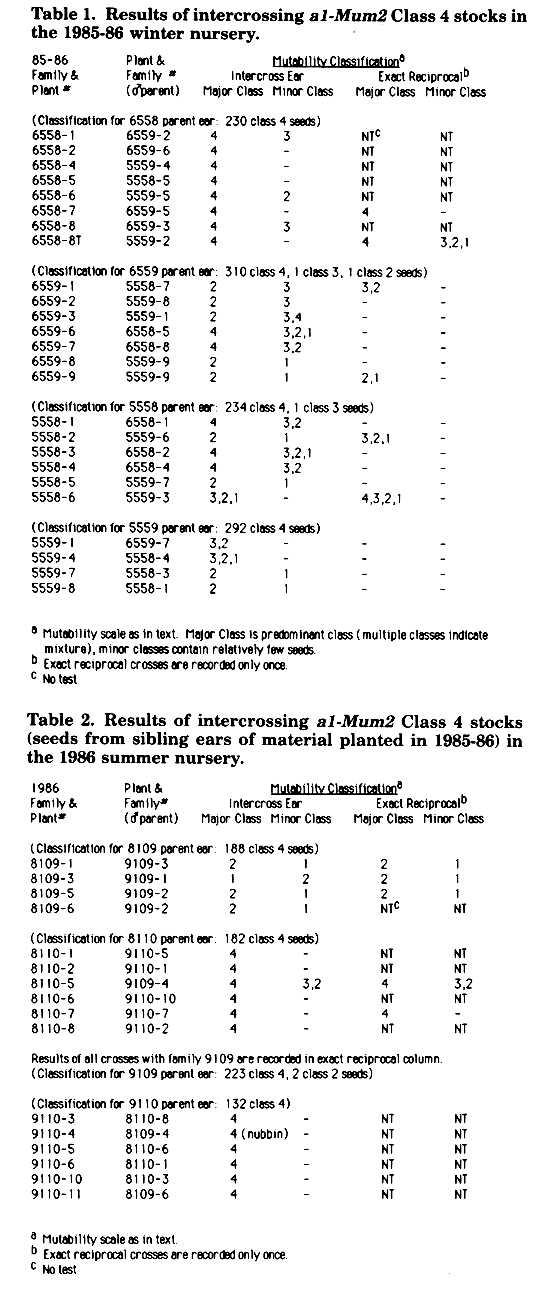
In the 1985-86 winter nursery, seeds from four of the ears from the summer crosses were sown (as families 5558, 5559, 6558, 6559) and the intercrossing was continued. The results of these crosses are given in Table 1. Although all ears from which seeds were selected for planting had only Class 4 or predominantly Class 4 seeds, intercrosses between three of the families resulted in ears which showed a marked loss in mutability. The one exception was family 6558. Whenever this family was involved in an intercross, the ears had only Class 4 or predominantly Class 4 seeds. The results were the same regardless of the direction of the cross. In all cases where the exact reciprocal crosses were made, the results were basically the same in both directions.
In the 1986 summer nursery, intercrosses between plants from the four additional 1985 ears were made. The results, shown in Table 2, are similar to those obtained in the winter nursery, in which seeds from sibling ears were planted. Again two types of families are seen. One type produces ears with primarily Class 4 seeds and the other type gives ears with predominantly low mutability seeds. Again, in crosses between high and low mutability lines, the high condition is dominant whether the high line is crossed as a male or female.
One of the 1985 ears, 6161-8/5151-3T had 237 Class 4, 64 Class 3 and 28 Class 2 seeds. Ten Class 2 seeds were planted from this ear. Eight plants in this family were either selfed and outcrossed to a1 sh2 or reciprocally crossed to a1 sh2. No mutable seeds were observed on any of the progeny ears.
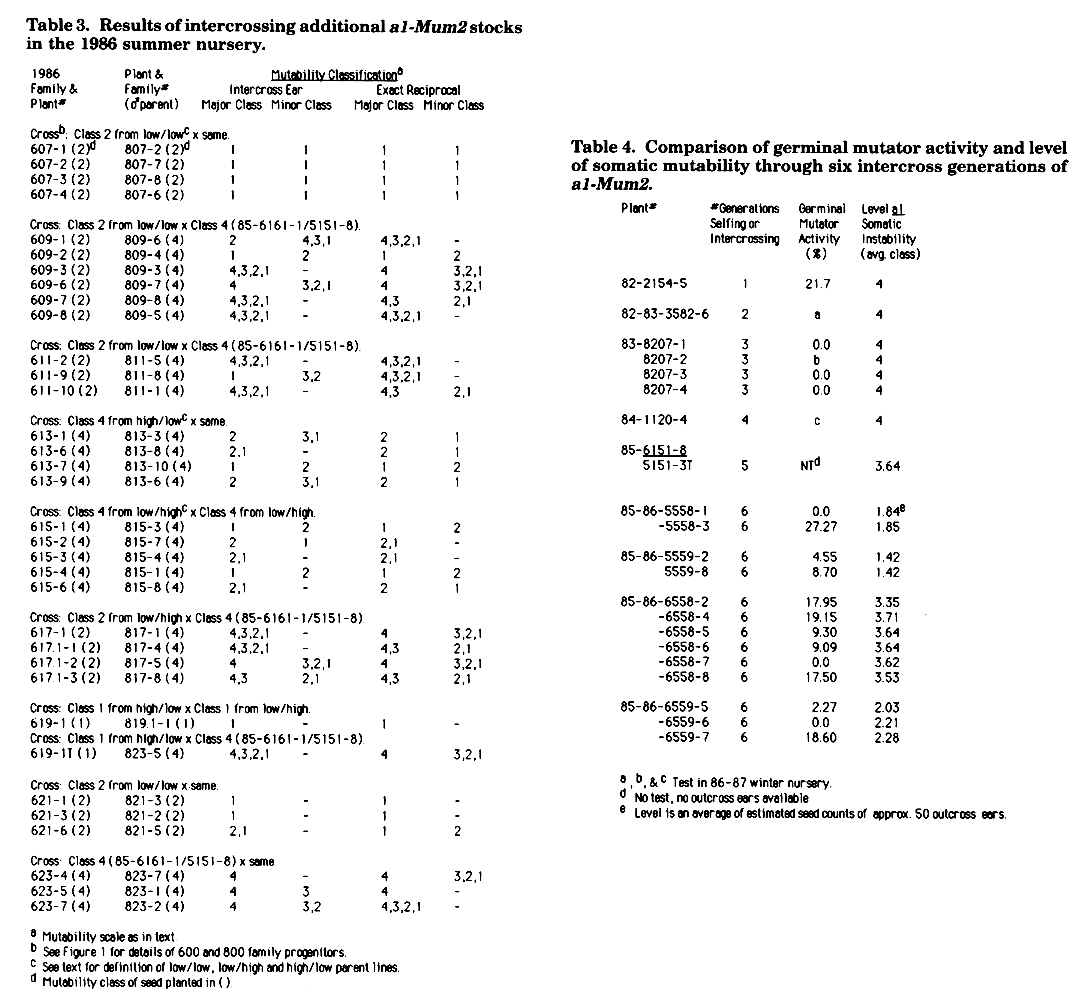
During the summer of 1986, seeds from the 1985-1986 winter nursery families 5558, 5559, 6558 and 6559 as well as some seed from the 1985 intercross ears were planted. The resulting plants were intercrossed reciprocally. Intercross results are reported in Table 3. All seeds used in these crosses were either 1985 class 4 seeds (as indicated) or derived from the 1985-86 intercrosses shown in Table 1. The following define the parents of the various crosses shown in Table 3: 1) Low/low-the result of intercrosses involving combinations of 85-86-5558, -5559, or -6559 families. 2) High/low-the result of intercrosses between family 85-86-6558 as a female times 85-86-5558, -5559 or -6559. 3) Low/high-the result of reciprocal crosses in which 85-86-6558 is the male parent. The specific family origin of the 86-600, and -800 families from Table 3 is shown in the pedigree in Figure 1.
When plants from Class 2 seeds from a low/low (607, 807) were intercrossed, the resultant progeny were without exception Class 1 stables. Similar results were obtained in exact reciprocal crosses.
Intercrosses between plants from Class 2 seeds from a low/low (609 and 611) and plants from a 1985 Class 4 (809 and 811) gave variable results. Seed mutability ranged from Class 4 to Class 1 with most ears segregating for Class 4 as a major class. Notable exceptions were ears from 609-1/809-6, 609-2/809-4 and 611-9/811-8 which gave primarily low seeds. Reciprocal crosses from these same plants showed higher mutability, however, all other sets of reciprocal crosses in these two families showed only a slight bias toward higher mutability when the Class 4 parent was the female.
Intercrosses between plants from Class 4 seeds from two high/low lines (613,813) gave only seeds of low mutability when crossed in either direction. Similarly, intercrosses between plants from Class 4 seeds from a low/high (615,815) gave seeds of low mutability. When plants from Class 2 seeds from a low/high (617,617.1) were intercrossed with plants from 1985 Class 4 seeds (817), all classes of mutability resulted on the progeny ears. Exact reciprocal crosses showed no outstanding reciprocal differences.
Intercrosses between two plants from Class 1 seeds, one from a high/low (619) and one from a low/high (819.1), produced only stable seeds when crossed in either direction. The tiller from 619-1 was crossed as a female with a plant from a 1985 Class 4 seed (823-5). This cross produced all classes of seeds, whereas the exact reciprocal cross produced predominantly Class 4 seeds with a few of each of the other classes. Intercrosses between plants from Class 2 seeds from a low/low (621,821) without exception resulted in ears with seeds of low mutability, or Class 1 stable seeds. The latter class was the predominant class on these ears.
The inbreeding of 1985 Class 4 plants was continued (623,823) with sibling seeds of those planted in the 1985-86 family 6558. Although some reduction in the level of somatic mutability was observed, the majority of the seeds produced were of high mutability. This was also the case for plants that were selfed (data not shown).
Levels of somatic mutability and germinal mutator activity are shown in Table 4 for individual plants from the pedigree of the stocks described in this report. An outcross of 82-2154-5 was selfed and seedling-tested for the production of new mutants. Of the outcross ears, 21.7% segregated for new mutations. After one generation of selfing and an intercross generation, 3 of 4 Class 4 siblings tested in the 83-8207 family had no germinal activity. Plants 83-8207-1 and -8207-2 were used in the production of the two Class 4 lines that were crossed in 1985 (see Figure 1). An outcross of 8207-2 has been planted and will be selfed in our 1986-87 winter nursery, then seedlings tested later this spring. This information is needed before we can fully interpret the data in this report. The first time we saw a significant reduction in the level of somatic mutability was in 1985. No outcrosses of the 1985 plants were made and thus germinal activity of these plants could not be determined. However, the 1985-86 families have been tested for germinal mutator activity. Five of the six plants from family 6558 which showed high somatic mutability, had germinal activity. The other outcross had no mutator activity. On the other hand, plants from the low families, 5558, 5559 and 6559, which when outcrossed gave mainly low seed mutability, showed no correlation between loss of somatic mutability and the presence or absence of germinal mutator activity. For example, outcrosses of 5558-1 and 5558-3 both resulted in seeds of low mutability, yet 5558-1 had no germinal activity and 5558-3 had germinal activity. Both 5559 siblings and 2 siblings in family 6559 showed such activity, while one did not.
A limited amount of molecular work on these lines has been initiated on selected plants in this pedigree. Table 5 summarizes information obtained to date as the result of Southern blot hybridization analyses. Individual plants from the pedigree in Figure 1 have been analyzed for Mu sizes, copy number, and modification of Mu element Hinfl sites. All plants tested contain both Mu1 and Mu1.7. The total copy number of these elements is approximately 15-20 per diploid genome. The copy number remains constant through the fifth generation of inbreeding. Extensive modification of the Mu elements does not occur until there is a loss of somatic mutability. This marked loss occurred in several of the 1985-6161/5151 crosses. When DNA from plants derived from Class 4, Class 3 and Class 2 seeds (85-86-6161-8/5151-3T) is analyzed for the presence of modification, there is a correlation between the amount of modified sequences and the relative level of somatic mutability. Although this has not been quantified, qualitatively, plants from Class 4, Class 3 and Class 2 seeds show increasing amounts of modified Mu elements relative to unmodified elements respectively. However, it is important to note that both modified and unmodified Mu sequences are present in all plants tested from this ear.
Germinal mutator activity and modification results for plants 82-2154-5 and 83-8207-1 indicate that in this material there is not a correlation between element modification and mutator activity. This may be further substantiated when the seedling test of 84-1120-4 is completed later this spring.
Plants 82-2154-5 and 83-8207-1 have the same number of Mu elements, none of which are modified, yet 82-2154-5 has germinal Mu activity while 83-8207-1 does not. Thus plants from seeds with high somatic mutability and possessing the same copy number of Mu elements do not necessarily have the same germinal activity. This lack of correlation of germinal activity with Mu copy number and somatic mutability would suggest that neither of these criteria are good predictors of Mu germinal activity. The results presented in Table 5 would suggest that the lack of modification also is not a good predictor of Mu germinal activity. The lack of any criteria that will predict germinal Mu activity suggests that the regulation of such activity is still unknown.
The following conclusions can be drawn from this work:
1) High somatic mutability can be retained after six generations of inbreeding.
2) However, in some sixth generation offspring, somatic mutability has been greatly diminished (low lines). This takes place suddenly in one generation.
3) Results from crosses between plants from high mutability seeds and those from low mutability seeds are the same, no matter what direction the cross is made. Thus there is no female effect observed.
4) When plants from Class 4 seeds from high/low and low/high crosses are crossed together, only stable or very low mutability seeds are observed in the progeny. Low/high and high/low do not behave differently in this regard. Again, no female effect is observed. In these crosses the low mutability state seems to prevail in both high/low or low/high hybrids because most progeny seeds are of low mutability.
5) Plants from seeds of different levels of mutability from the same ear show a progressive degree of modification of the Mu elements present that is inversely proportional to the level of somatic mutability.
6) There is little or no correlation of Mu germinal activity with the total Mu element copy number, the Mu element modification or somatic mutability.
Table 1. Results of intercrossing a]-Mum2 Class 4 stocks in the 1985-86 winter nursery.
Table 2. Results of intercrossing a1-Mum2 Class 4 stocks (seeds from sibling ears of material planted in 1985-86) in the 1986 summer nursery.
Table 3. Results of intercrossing additional a1-Mum2 stocks in the 1986 summer nursery.
Table 4. Comparison of germinal mutator activity and level of somatic mutability through six intercross generations of a1-Mum2.
Table 5. Summary of Southern blot hybridization results to date.
Bradley A. Roth and Donald S. Robertson
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}