All Mu-induced mutable aleurone genes found to date in our studies with Mu have had a very characteristic spotting pattern of small spots, indicative of late events. Although there is some variation from mutant to mutant in the size of the spots, they are nonetheless always the result of late events. Large, irregular pigmented spots, which would be expected from early events, are rarely observed.
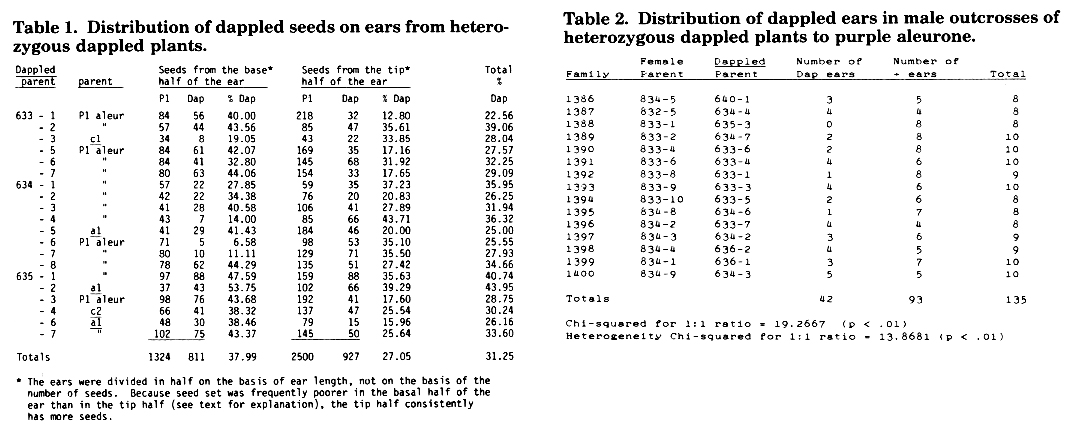
From our 1979 a2 bt x purple Mutator isolation plot, we recovered many seeds that had sectors of purple and yellow aleurone. These seeds did not have the typical Mu aleurone mutable pattern, but instead had varying sectors of purple and yellow. A couple hundred such seeds with different amounts of sectoring were planted in 1982 and self pollinated. All but a very few segregated for purple Bt and yellow bt seeds. Thus, the original sectoring was probably due to chromosome loss or somatic mutation within the endosperm. However, one plant from a sectored seed segregated for a mutable aleurone pattern we have called dappled (Dap). This mutable pattern is not typical of previous Mu-induced aleurone mutants. Dap seeds are yellow with sectors of purple tissue of variable sizes and shapes (Figures 1 and 2). Dap seeds are frequently smaller than their purple sibling seeds, and are sometimes extremely defective (Figure 2). There appears to be an imperfect correlation of this defective phenotype with the extent of yellow areas on the seeds. The greater the amount of yellow tissue, the smaller the seed. Frequently, the region of the ear that matures first will have a higher concentration of dappled seeds than the tip of the ear (Table 1); these dappled seeds have more yellow tissue than those at the tip of the ear. In most cases where this pattern is found, there is irregular seed set in this region of the ear of the type seen on a pollinated ear in which the silks have passed the prime time for pollinating. However, this is probably not the explanation for most of the ears observed in our tests because most were pollinated within a couple of days of first silking. Because seeds with very little purple tissue are very defective, it may be that seeds without any purple sectors abort. Such seeds would be more likely in this region of the ear where the dappled expression is more intense.
It was first assumed that dappled was a mutable allele of a2 because it was observed in an a2 bt x purple Mu cross. However, to rule out the possibility that another aleurone locus was involved, it was tested against the aleurone testers for a1, a2, c1, c2, and r loci. These tests were made reciprocally. At the same time, Dap stocks also were crossed reciprocally to purple aleurone lines. The plants that were used in these crosses were from Dap seeds. All crosses in which Dap plants were used as males gave nothing but purple seeds. (Two sectored seeds from male outcrosses to a2 testers were found, but when they were planted and test crossed, they turned out not to be dappled.) However, all crosses where Dap plants were used as females segregated for Dap seeds, even the crosses with purple aleurone. Thus, Dap seems to behave as a dominant trait in plants crossed as females.
Although dappled is not expressed in male outcrosses, it is male transmissible, though in a reduced frequency. Last summer, we planted 10 seeds from each of 15 different male outcrosses of dappled to purple aleurone, and selfed these plants (Table 2). Had male transmission been complete, half of the plants would have segregated for dappled seeds (the male parents were heterozygous). The reduced frequency of dappled ears observed (31%) was significantly lower than 50% (p < .01). This summer, we will test a larger number of crosses in which Dap plants are used as males, to obtain a better estimate of its rate of transmission through the male.
As mentioned above, expression of dappled in female outcrosses is reduced from the expected frequency of 50% (Table 1). Furthermore, there is a higher frequency of Dap kernels at the base of the ear than at the tip. These observations have two possible explanations: (1) Female transmission of Dap is reduced, and (2) for whatever developmental reasons, Dap is not expressed as extremely at the tip of the ear as it is at the base; thus, more of the kernels at the tip could be heterozygous, but the trait isn't being expressed. We will test both of these possibilities this summer by growing purple kernels from both the tip and base regions of these ears, and seeing if they carry dappled.
All plants from dappled seeds so far studied have been heterozygous, even those arising from seed from selfed Dap plants, which suggests that the gene is lethal in the homozygous condition. If further tests bear this out, there could be a very interesting dosage pattern regulating the expression of this gene (i.e., + + + and Dap + + = purple aleurone, Dap Dap + = mutant aleurone, Dap Dap Dap = lethal). On the other hand, the difference in expression between male and female outcrosses could be due to male vs. female transmission effects. We will investigate these possibilities at the earliest opportunity, and welcome suggestions for further experiments.
Under the dissecting microscope, the purple areas of dappled seeds appear to be raised above the yellow sectors. If the pericarp is peeled from the seed, the cellular structure in the central portions of the yellow areas appears amorphous and irregular. The purple sectors have normal-appearing aleurone, and the portions of the yellow areas that are immediately adjacent to purple sectors are normal, too.
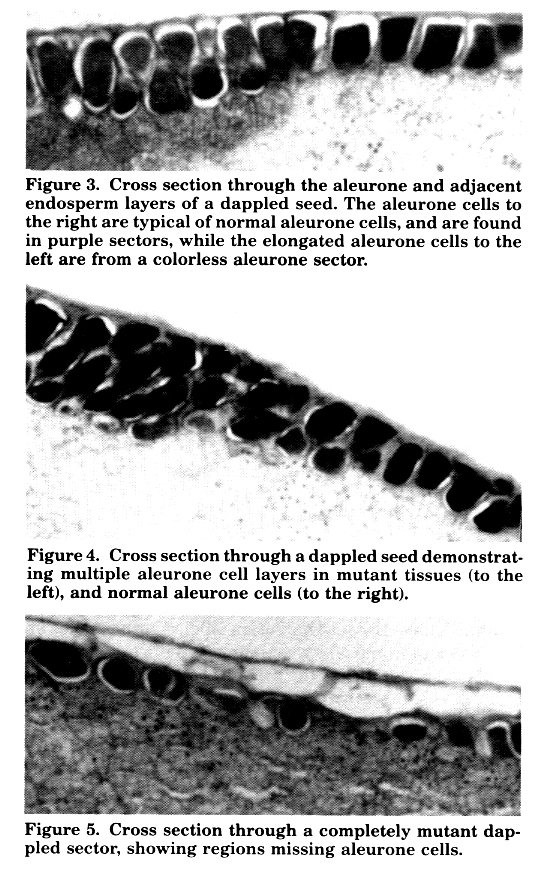
Cross sections through the aleurone and adjacent endosperm of imbibed Dap seeds (paraffin-sectioned, stained with safranin and counter-stained with chlorazol black-prepared by PSS) reveal normal-appearing, intensely staining aleurone cells in the purple sectors (Figures 3-5). In the colorless sectors, several patterns of aleurone cell morphology are observed. In some instances, the aleurone cells are lighter staining, and elongated (about twice as long in the direction perpendicular to the seed surface as they are in directions parallel to the surface-Figure 3). In other preparations, the aleurone in the colorless sectors consists of several layers of small, irregularly-packed cells (Figure 4). In still other preparations, the colorless sectors have an aleurone consisting of a few "normal" aleurone cells irregularly interspersed with what appear to be starchy endosperm cells similar to the ones which normally underlie the aleurone (Figure 5).
Although some of these cell patterns may be the result of artifacts induced by our cytological technique, which we are still trying to perfect, it is nonetheless apparent that the Dap mutant is altering the normal development of the aleurone cells, and since the mutant cells are colorless, these altered cells have lost the ability to synthesize anthocyanin pigments. This alteration has more severe effects than just altering pigment synthesis as evidenced by the very defective state of seeds that are predominately yellow with very little normal aleurone tissue, and the total absence of completely yellow seeds. Whatever aleurone function is interrupted by this mutation, it appears to be essential for normal seed development. Thus, Dap is undoubtedly a developmental mutant.
Dappled seems to be a dominant mutable gene affecting the development of the aleurone. It has reduced transmission through the male (and possibly through the female as well), and may be dose-dependent in its expression.
Two additional mutants with aleurone patterns that superficially resemble dappled have been found in Mu aleurone color stocks. One arose from a cross of a purple Mu plant with an a1 sh2 tester. This mutant is similar in both aleurone phenotype and transmission. It is not allelic to a1, and the sectored seeds are only found in female crosses and not in male crosses. It is probably allelic to Dap, but since we have not yet demonstrated this conclusively, we are calling this mutant Dap*-3349. The second mutant occurred in a Mu-induced bronze-1 mutable stock (bz1-Mum8). Plants from the sectored seeds were twisted, crinkly-leaf dwarfs with tassels having anthers that dehisce poorly. We are currently in the process of further characterizing this mutant (Dap*-6143), which appears to be expressed in both male and female outcrosses.
Figure 1. Close-up of seeds near the base of a dappled ear. Note that there are large and small Dap seeds, and that there appears to be an inverse relationship between seed size and the amount of mutant tissue present. Very small, defective Dap seeds are not seen in this picture because they are usually overgrown by more fully developed seeds.
Figure 2. Close-up of dappled seeds.
Table 1. Distribution of dappled seeds on ears from heterozygous dappled plants.
Table 2. Distribution of dappled ears in male outcrosses of heterozygous dappled plants to purple aleurone.
Figure 3. Cross section through the aleurone and adjacent endosperm layers of a dappled seed. The aleurone cells to the right are typical of normal aleurone cells, and are found in purple sectors, while the elongated aleurone cells to the left are from a colorless aleurone sector.
Figure 4. Cross section through a dappled seed demonstrating multiple aleurone cell layers in mutant tissues (to the left), and normal alettrone cells (to the right).
Figure 5. Cross section through a completely mutant dappled sector, showing regions missing aleurone cells.
Philip S. Stinard and Donald S. Robertson
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}