The a-m-4 mutable allele of the anthocyanin locus, A1, originated from a wildtype A1 allele in a plant carrying the controlling elements Dissociation (Ds) and Activator (Ac) (B. McClintock, Genetics 38:579-599, 1953). Ac controls transposition both of itself and of Ds causing both insertional inactivation of genes and subsequent reversion (i.e., a mutable phenotype). Since a-m-4 mutability is controlled by, but unlinked to Ac, it is thought that the a-m-4 mutation resulted from a Ds insertion. Ds elements isolated so far fall into two classes based on their degree of homology with the Ac element. We were interested in identifying the Ds element thought to be present in a-m-4 and in determining its position relative to the A1 gene.
Molecular analysis of a-m-4 was facilitated by the availability of a cloned A1 gene and two cloned Ds elements for use as hybridization probes. To isolate a-m-4 clones we constructed a genomic library in lambda EMBL4 from an a-m-4 homozygous plant (no Ac elements present in the line, seed provided by M.G. Neuffer) and screened the library with an A1 probe (C. O'Reilly et al., EMBO J. 4:877-882, 1985). We then screened A1-homologous clones with the Ds probes derived from two different Ds elements; Ds1, which is nonhomologous to Ac (isolated from the wx-m1 allele, and provided by S. Dellaporta; S.R. Wessler et al., EMBO J. 5:2427-2432, 1986), and Ds-5933, a deletion derivative of Ac (isolated from the sh-m5933 allele, and provided by H.P. Doring; H.P. Doring et al., Nature 307:127-131, 1984). Three A1-homologous clones hybridized to the Ds1 probe but none hybridized to the Ds-5933 probe. Our preliminary results suggest that the mutability of a1 gene expression in the a-m-4 allele isdue to a Ds1-homologous element.
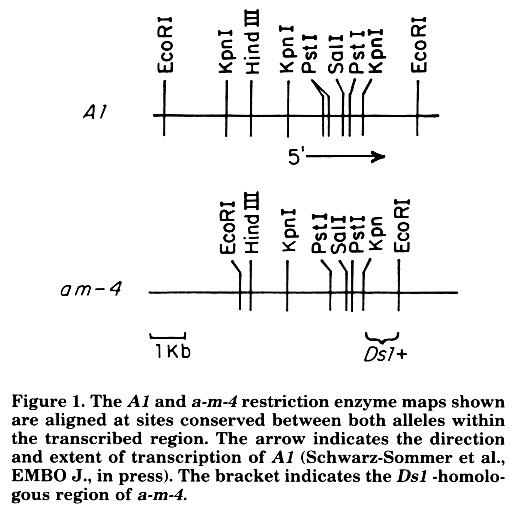
We constructed a restriction enzyme map of a representative a-m-4 clone and determined the region of Ds1 homology by Southern analysis and hybridization to the Ds1 probe (Fig. 1). A comparison of the preliminary a-m-4
data with a wildtype A1 restriction enzyme map showing the region of A1 transcription indicates that the Ds1 homology is located toward the 3' end of the gene. The Ds1-homologous region is 3' of other transposons at A1: those reported in Zs. Schwarz-Sommer et al. (EMBO J., in press), the Mul element in a1-Mum2 (5' of the A1 transcription unit), the two rDt insertions in a-m-1:Cache and a1 (in exons 3 and 4), and the rMrh insertion in a-mrh (also in exon 4).
Differences in EcoRI sites between a-m-4 and A1 (shown in Fig. 1) may be restriction fragment length polymorphisms resulting from minor sequence alterations or from more extensive differences. Such EcoRI polymorphisms are common between A1 alleles (for example, A1 alleles reported in Zs. Schwarz-Sommer et al. EMBO J., in press; and a1, a-m-1:Cache alleles which were cloned on 10kb EcoRI fragments).
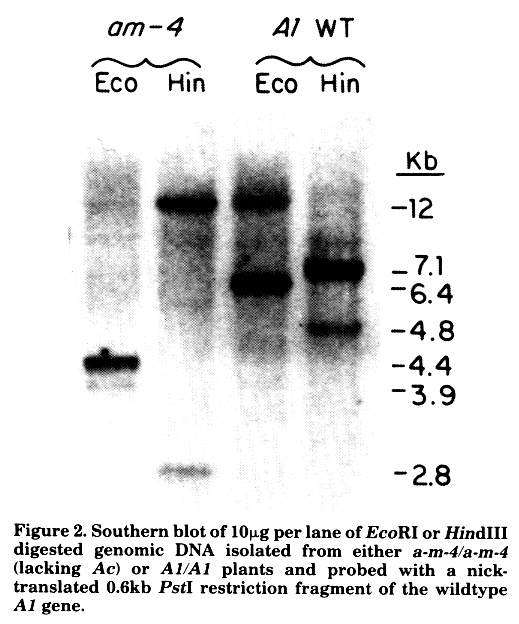
A genomic Southern blot of DNA from a-m-4 or A1 homozygous plants probed with a fragment of the wildtype A1 gene is shown in Figure 2. The sizes of the major hybridizing bands agree with the restriction maps constructed from a-m-4 and A1 clones. Maize genomic Southern blots probed with wildtype A1 sequences often result in one major hybridizing band and a second minor band, which may be either smaller or larger than the major band. As shown in Figure 2, this is the case with DNA prepared not only from plants carrying mutable alleles of A1, such as a-m-4, but also from those carrying a wildtype A1 allele. This might result from the presence of a second locus that shares incomplete sequence homology with A1. Alternatively, the minor bands may represent somatic rearrangements of A1 sequence in certain cells even when known transposons are not active in the genome. One A1-homologous clone isolated from the a-m-4 library seems to have the same structure as the minor bands seen on the genomic Southern of a-m-4/a-m-4 DNA (Fig. 2). We will analyze both the Ds1-hybridizing genomic a-m-4 clone and this second type of genomic clone in order to determine their relationship.
J.J. Sorrentino and N.S. Shepherd
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}