Lesion expression in Les1 homozygotes is earlier, more uniform, and more severe than in Les1 heterozygotes. Consequently, it is preferable to use homozygous material for biochemical and physiological investigations of Les1 expression. One can actually score Les1 homozygous seedlings before lesion expression is initiated with the aid of the closely linked recessive marker wt1 (white-tipped first and second leaves) (Hoisington, MNL 60:51). Such marked stocks are generated simply by selfing or intercrossing + +/Les1 wt1 lines. (Les1/Les1 plants die as seedlings in the field.) The expected ratio of 1 wt1:3 Wt1 seedlings was not found, however, in most of the stocks generated in this manner.
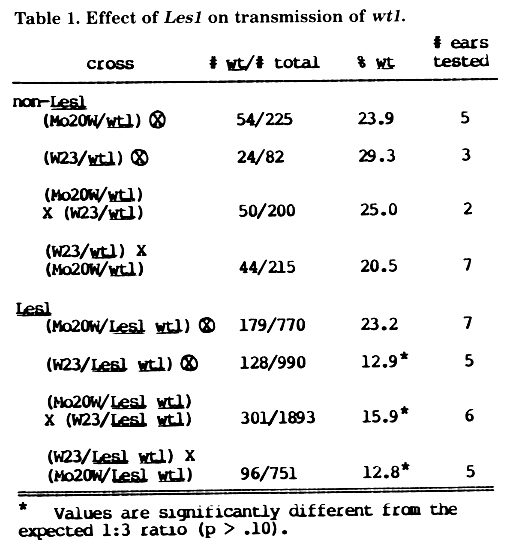
Table 1 lists the percentages of wt1 seedlings observed for several different types of + +/Les1 wt1 x + +/Les1 wt1 crosses. The +/wt1 x +/wt1 crosses used as controls were made with segregating +/wt1 (non-Les1) siblings. The two F1 families used in these crosses had as their male parent the same + wt1/Les1 wt1 individual. In all crosses 25% of the progeny are expected to be wt1.
The data indicate that all crosses involving W23 and the Les1 wt1 chromosome have a striking reduction in the number of Les1 wt1 progeny. Germination frequencies for all families were between 97% and 99%. Semi-sterility was not evident on any of the ears from + +/Les1 wt1 plants. Pollen abortion in the + +/Les1 wt1 plants was not examined. Sufficient data on the reciprocal crosses between + +/Les1 wt1 and +/wt1 plants are not yet available.
Under-representation of Les1 wt1 seedlings cannot simply be due to poor expression of the wt1 trait for two reasons. First, very few of the non-wt seedlings gave lesion expression characteristic of Les1 homozygotes. This type of lesion expression would be expected among the Wt1/class if the Les1 wt1 chromosome was transmitted at a normal frequency but wt1 was poorly expressed. Second, outcrosses of the original recombinant + wt1/Les1 wt1 individual (recovered by D. Hoisington), and of the + +/Les1 wt1 F1 individuals, onto various inbred lines resulted in a lower than expected frequency of +/Les1 plants in the progeny. In these crosses Les1 gametes (microspores) were transmitted at a frequency of 31% (32/103) from the original + wt1/Les1 wt1 individual, 38%, (29/77) from several Mo20W/Les1 wt1 F1 individuals and 18% (40/223) from several W23/Les1 wt1 F1 individuals. Similar crosses with + /Les1 plants give the expected transmission frequency of 50% for Les1 gametes (Neuffer & Calvert, J. Heredity 66:265).
Further tests are needed to explain why the lower transmission of Les1 gametes that was observed following outcrossing of the Mo20W/Les1 wt1 family was not observed following its selfing. If Les1 is directly involved in the reduction of Les1 wt1 gametes, however, then another interesting effect of the Les1 mutation has come to light.
Craig Echt
Return to the MNL 61 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}