We have recently isolated and positively identified a genomic clone of the gene for sucrose synthetase-2 (Ss2) in maize. The clone was isolated by exploiting the partial homology between Shrunken-1 (Sh1) and Ss2. A BamHI genomic library was constructed in EMBL 3 lambda phage using DNA from a single Black Mexican sweet corn plant kindly provided by Paul Still and Prem Chourey The library, consisting of approximately 300,000 plaques, was screened with a 3400 bp BgIII fragment from the Sh1 clone p17.6 (Sheldon et al., Mol. Gen. Genet. 190:421-426, 1983; Zack et al., in press). The probe begins in intron 2 of Sh1 and ends in exon 14.
Nine strongly hybridizing plaques were isolated and were subjected to analysis. Of these clones, eight contained inserts that corresponded to either of two wild type Sh1 alleles as judged by restriction mapping. The original plant from which these clones were derived was apparently heterozygous at Sh1. One of these alleles appeared to be identical to our original Sh clone, p17.6 (except that the new clones are BamHI fragments and p17.6 is an EcoRI fragment), while the second allele exhibited small but interesting differences in its restriction patterns.
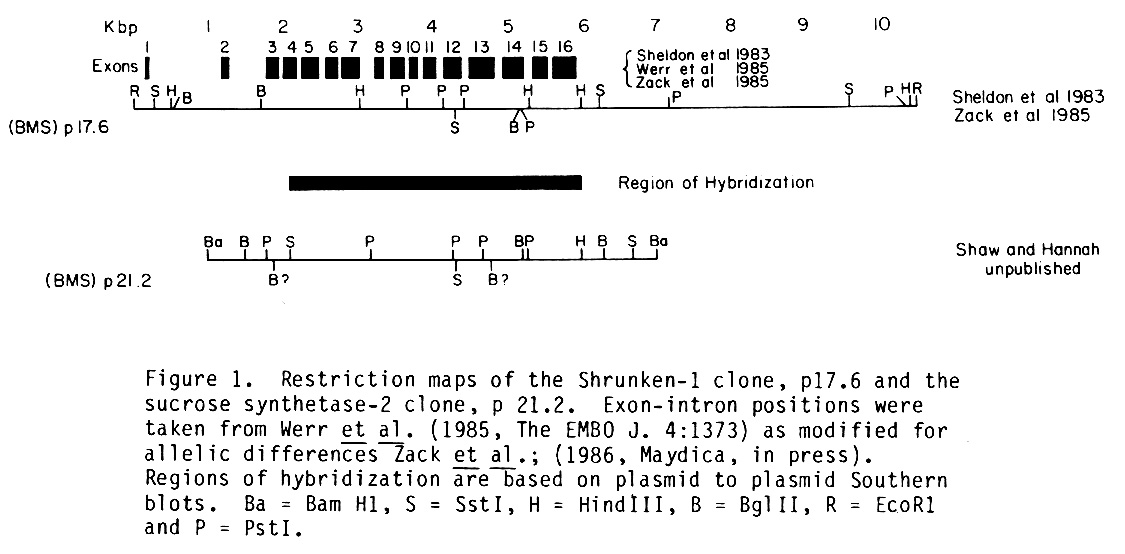
The ninth clone (designated lambda 21.2) differed markedly from the others. BamHI digestion of lambda 21.2 revealed three inserted fragments of DNA. Only one of these, a 6.6 kb BamHI fragment, hybridized to the 3400 bp BgIII fragment of p17.6. The origin of the other two fragments is unclear. They may be random fragments of maize DNA which ligated to the 6.6 kb fragment during cloning, or the original BamHI digestion used to construct the library may have generated incomplete cleavage products. The 6.6 kb fragment was subcloned and is designated p21.2. Restriction maps of p21.2 and p17.6 are shown in Figure 1. The restriction maps of p21.2 and p17.6 bear only a modest resemblance. However, the two clones share substantial regions of homology as indicated by plasmid to plasmid Southern blot hybridization (solid bar). Thus, we had isolated a genomic clone with partial homology to Sh1 from a library in which we had already identified both of the Sh1 alleles.
We considered several possible explanations for the origin of p21.2: (1) p21.2 arose as a cloning artifact via an in vitro rearrangement of Sh1 sequences or rearrangement of Sh sequences during plant growth. (2) p21.2 represents an inactive copy of the Sh1 locus (i.e. a pseudogene). (3) p21.2 is a clone containing all or part of the sucrose synthetase-2 gene.
To test the first possibility we performed Southern blot analysis of DNA isolated from wild type and sh1 bz-m-4 plants (sh1 bz-m-4 lacks Sh1 sequences). p21.2 sequences were clearly present in both wild type and sh1 bz-m-4 stocks. Thus, p21.2 did not arise from an in vitro or in vivo rearrangement of Sh1 sequences. Interestingly, however, the restriction fragment banding pattern observed in the two genotypes differed, indicating that a polymorphism exists between these genotypes. In principle, these stocks should be isogenic except in the region of Sh1 on chromosome 9. This suggested to us that the p21.2 sequence may also be located on chromosome 9. Further mapping experiments have corroborated this (McCarty et al., this issue below).
In order to determine whether p21.2 represented an active gene, we examined polyA-RNA isolated from developing seeds of wild type and sh1 bz-m-4 deletion stocks by Northern blot analysis. As expected p17.6, the Sh clone, hybridized strongly to an abundant 2.75 kb mRNA which was present in the wild type but absent in the sh1 deletion stock. A second weakly hybridizing mRNA of slightly greater molecular weight could be identified in the deletion stock. p21.2 hybridized strongly to mRNA that was present in equal abundance in both wild type and deletion stocks. This mRNA is identical in size to the transcript that showed weak hybridization to Sh1. Weak hybridization of p21.2 to the Sh1 transcript in the wild type was also evident. Therefore p21.2 represents an active gene distinct from Sh1, and this gene is most probably sucrose synthetase-2. The slightly higher molecular weight of the putative Ss2 transcript is consistent with the observation of S. McCormick et al. (Mol. Gen. Genet. 187:494, 1982).
To positively identify the clone, we purified the p21.2 transcript from polyA-RNA extracted from sh1 bz-m-4 by hybrid-selection with p21.2 DNA covalently linked to a solid matrix. When translated in vitro in a rabbit reticulocyte lysate system, the hybrid-select mRNA produced a single translation product with a molecular weight of 88,000 daltons, the size of the sucrose synthetase-2 subunit. Furthermore, using Sh1 anti-serum (Sh1 anti-serum generously provided by Paul Still and Prem Chourey will cross-react with sucrose synthetase-2) we can specifically immunoprecipitate the hybrid-select translation product. Thus, a p21.2 is a genomic clone containing the gene for sucrose synthetase-2.
From sequencing experiments we have found homology between p21.2 and p17.6 beginning at the SstI site in exon 12 of Sh1 and extending 3' into intron 12. Some 20 bp into intron 12 homology breaks down but resumes several base pairs before the intron 12-exon 13 border. It would appear that sequence organization of Sh1 and Ss2 is similar, at least in the vicinity of intron 12.
D.R. McCarty, J.R. Shaw and L.C. Hannah
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}