Zygomeres are units that are responsible for chromosome pairing. Their existence is, as yet, hypothetical. The length of the synaptonemal complex (SC) is very much shorter than the length of the DNA. In corn, for example, the SC:DNA ratio is 1.5:10,000 (Gillies). This implies that there is a special class of DNA responsible for chromosome pairing, because a random sample of DNA from each chromosome would have a very low probability of being homologous. These specialized DNA segments will be called zygomeres.
One way to establish the existence of zygomeres is to study preferential pairing in trisomes that have two homologous chromosomes and one odd homoeologous chromosome. Preferential pairing can be determined by its effects on genetic ratios as shown in Table 1. When the trisomes are used as the pollen parent only the monosomic gametes need to be considered because disomic gametes function in fertilization only very rarely. If only homologous pairing occurs, then only homogenetic bivalents (BU-A) will be formed, and all the gametes will be g. If the preferential pairing factor (p) has a value, then the random ratio of 1 G:2 g will change. The "frying pan" trivalents (T-3A and T-3B) behave like the bivalent and univalent types (BU-A and BU-B) because the chromosomes forming the "pan" will disjoin and go to opposite poles. This assumes that the chromosomes are reductional (GG or gg) instead of equational (Gg) as the result of crossing over. The chain trivalents (T-1A and T-1B) will not give a 1 G:2 g ratio if the frequency of alternate segregation is greater than the random value of 1/3. The frequency and placement of chiasmata are probably affected by the environment, and different relative frequencies of pairing configurations will result with concomitant changes in genetic ratios. Therefore, the expression of preferential pairing is somewhat variable.
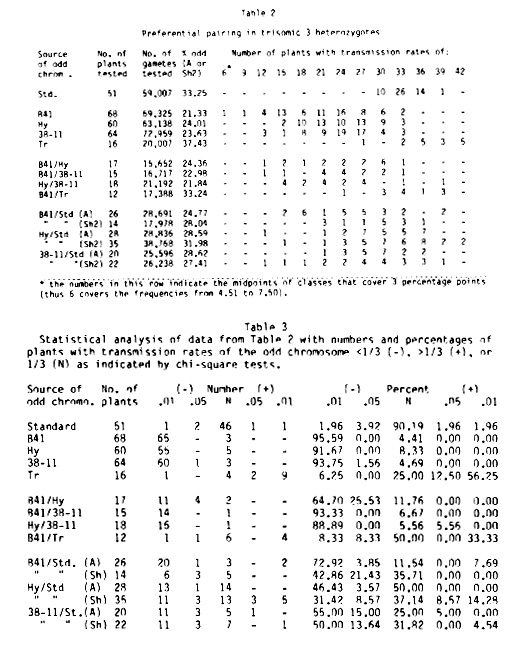
Trisome 3 plants (a Sh2/a Sh2/a sh2 or a Sh2/a sh2/a sh2) with all standard chromosome 3's have been crossed with many sources of possible homoeologous chromosomes that are marked with A Sh2. The progeny of these crosses (A Sh2/a Sh2/a Sh2, A Sh2/a Sh2/a sh2, or A Sh2/a sh2/a sh2) are crossed as the male with a sh2/a sh2 testers. Four inbred lines (B41, Hy, 38-11, and Tr) whose chromosome 3's produced a strong preferential pairing response have been selected for an intensive study. Hybrids between these lines and hybrids between them and the standard have been used in crosses with the standard trisome 3 line. Recombinant chromosome 3's with different patterns of zygomeres were anticipated. The results are shown in Table 2, statistically evaluated in Table 3.
If the data in Table 2 are plotted graphically, in most cases a normal curve is approximated. In some cases, such as B41, a bimodal distribution is found. In all cases the variability of preferential pairing allows for the upper tail of these normal curves to extend over the random pairing point (33.3%) and gives false readings for normality. The means of transmission rates do not fluctuate significantly from year to year.
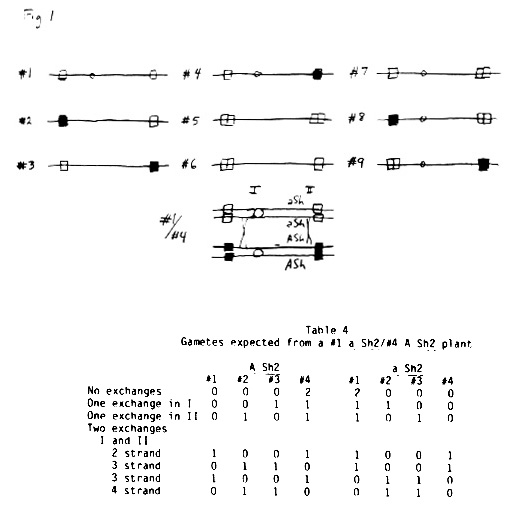
The data in Table 2 can be explained by a simple model. It is assumed that chromosome pairing is initiated by two zygomeres, one each at or near the ends of the chromosome. There are nine possible patterns, as shown in Figure 1. The standard chromosome 3 is #1; the B41, Hy, and 38-11 chromosome Ts are #2, #3, or #4, and the Tr chromosome 3 is #5. The zygomeres indicated by the black squares are non-homologous to the standard white square zygomeres. The Tr zygomeres are thought to be homologous to the standard zygomeres, but are stronger. A Tr zygomere pairs more readily with a standard zygomere than do two standard zygomeres with each other. It is hypothesized that zygomeres consist of many tandemly repeated units and the Tr chromosomes have more units than the standard. Thus it gives p a negative value and the ratio is >1 G:<2 g (See Table 1).
The results from using hybrids B41/Hy, B41/38-11, and Hy/38-11 as the source of the odd chromosome indicate that the zygomere patterns of the hybrids are #244, #344, or #444. They are not #243 because crossing over would produce about 1/2 #1 chromosomes, assuming the zygomeres would show 50% recombination if they are at the ends of the chromosomes. The highest numbers of apparent #1 chromosomes, showing normal (33%) transmission, are from B41/Hy, where there are 11.76%. One or both of these chromosome pairs must be #4. Where the hybrid B41/Tr was used it may be seen that data, while limited, indicate segregation for <1/3 and >1/3 transmission of A, along with 1/3 transmission cases. Crossing over should produce #8 and #9 zygomere patterns which would be equivalent to #1 if these zygomeres acted independently to cancel out each other's effects.
The results from using the inbred line/standard hybrids as the source of the odd chromosome are quite instructive. If the Hy/Std is #241 we would expect an equal frequency of A Sh2 #1, A Sh2 #2, a Sh2 #1, and a Sh2 #2 chromosomes following crossing over. The data show that this appears to be true, although there is an unexplainable enhancement of pairing affinity in 22.85% of the a Sh2 chromosomes.
The other two inbred line/standard hybrids (B41/Std and 38-11/Std) are probably #4/#1. The consequences of crossing over in this case are shown in Table 4. If there are two exchanges the frequencies of A Sh2 and a Sh2 chromosomes with patterns of #1, #2, #3, and #4 are all 1/4. The chromosomes #2, #3, and #4 will give preferential pairing. The data are in fairly close agreement.
The mapping of zygomeres, like any gene, should be possible although very difficult.
Table 4. Gametes expected from a #1 a Sh2/#4 A Sh2 plant
G. G. Doyle
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}