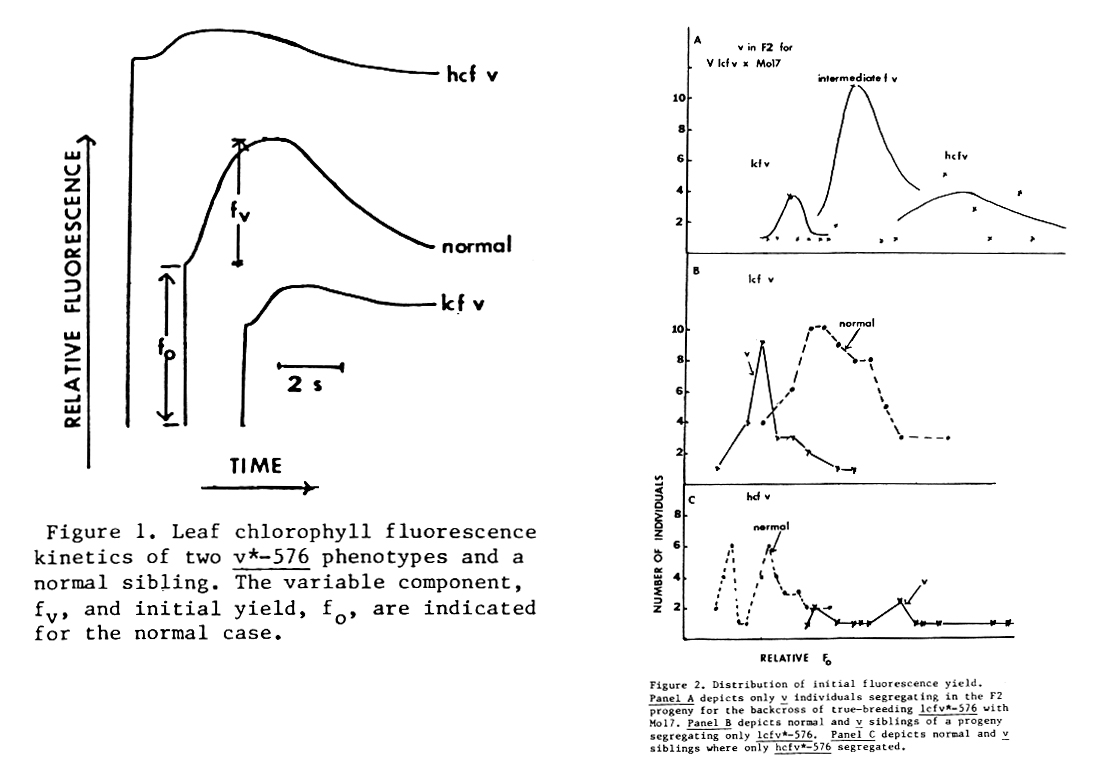
The nuclear mutation v*-576 was found by M.G. Neuffer in a M2 progeny derived from pollination with ethylmethylsulfonate-treated pollen. When crosses were made between inbred Mo17 and M2 material segregating v*-576, I found that most F2 progenies segregated two virescent phenotypes. A few F2 progenies segregated only one v-trait. I could distinguish the two v-phenotypes by noninvasive measurement of leaf chlorophyll fluorescence induction kinetics: one exhibited a high initial yield of fluorescence (hcf), the second, a lower than normal yield (lcf). Additionally, the ratio of the variable yield fluorescence component (fv) to the initial yield (fo) is greatly reduced for hcf-v but not lcf-v (see Figure 1). In both cases, abnormal fluorescence was developmentally conditional and observed only in partially, as opposed to completely, greened leaves. We have shown that hcf v*-576 is associated with early assembly of the major chlorophyll light harvesting complex (LHCII) and lcf v*-576 with late assembly of LHCII (Polacco et al., Plant Physiol. 77:795-800,1985). Normally, LHCII-assembly is a late event of thylakoid differentiation. Both v*-576 traits are allelic to v*-424 (MGNL 1985), a mutation similarly derived by M.G. Neuffer, which has continued to segregate only the hcf-v phenotype after four backcrosses to Mo17.
Either modifying loci or mutability at v*-576 are possible explanations for the two v*-576 phenotypes. My recent data indicate a modifying locus, Mof* (modifier of fluorescence), has been unmasked by v*-576. I have crossed Mo17 with individuals of progenies segregating only one v*-576 phenotype where at least eight, self-fertilized siblings, heterozygous for v*-576, were found to segregate only the same v-trait. I have only analyzed crosses involving the lcf-v trait. Both lcf-v and hcf-v individuals appear in F2 progenies derived from two F1 generations. A large class of v-individuals with an intermediate fluorescence pattern predominates. This class has normal or somewhat elevated initial fluorescence yield and a significant fv component. A histogram of the initial fluorescence yield data for palegreen v-leaf tissue shows a continuum of values that fit three superimposed, bell-shaped curves. The areas bounded by the curves approximate a 1:2:1 ratio for lcf:intermediate:hcf fluorescence yields (see Figure 2). Genetic analysis of the intermediate fluorescence yield class is required to confirm the 1:2:1 segregation. Typically, there is some overlap of analogous fluorescence distribution curves for normal and v siblings taken from progenies segregating only lcf v*-576 or hcf v*-576. The overlap probably reflects the conditional character of the phenotypes. I tentatively conclude that two variant, codominant alleles at an unlinked locus (Mof*) are involved, that Mo17 carries the allele responsible for the hcf-v trait (Mof*-1) and that the M2 stock carried both alleles (Mof*-1, Mof*-2). I am currently testing whether the putative Mof*-2 allele responsible for lcf-v can modify the hcf-v trait of other mutations allelic to v*-576.
Mary Polacco
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}