In higher plants it has been demonstrated that gametophytic traits are to some extent under the control of genes which are also expressed in the sporophyte (genetic overlap). Therefore gametophytic selection can lead to correlated sporophytic responses which can involve agronomic traits, too (D.L. Mulcahy, Science 171:1155, 1971; Ottaviano et al., TAG 6:249, 1982).
As part of current research efforts aimed at assessing the real value of this selection in applied breeding programs, a recurrent gametophytic selection was implemented in maize. The preliminary evaluation of both direct and correlated responses is presented herein.
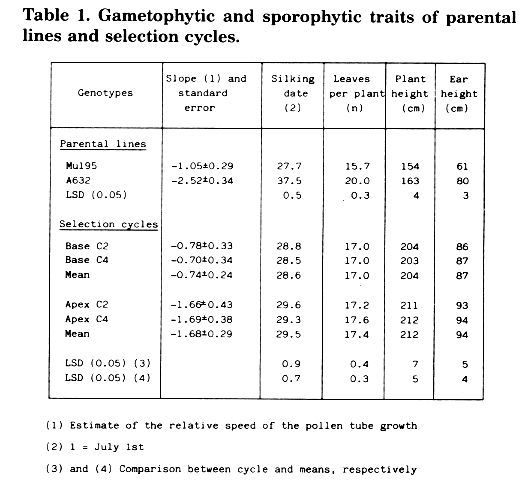
Gametophytic selection in maize is based on the silk length increase from the ear apex downward, which allows the pollen tubes with a higher growth rate to fertilize more frequently at the ear base than at the apex. As a breeding population we used the F2 of the cross between the inbred lines Mu195 and A632, which show substantial differences for both gametophytic and sporophytic traits (Table 1). One hundred full-sib crosses were made between random F2 plants. Their ears were shelled by taking 25 kernels from the base and as many from the apex, which were bulked according to the ear segment they were taken from, thus obtaining the Base C1 and the Apex C1. In subsequent years, while 100 full-sib crosses were made within each cycle as previously indicated, their ears were shelled by taking 25 kernels at the base only if derived from Base cycles, or at the apex only if derived from Apex cycles.
To evaluate the responses to selection, the 2nd and 4th Base and Apex cycles were compared together with the parental lines in a field trial with 4 replications. The gametophytic trait, i.e. the pollen tube growth rate, was measured in vivo by means of the pollen mixture technique (E. Ottaviano et al., TAG 63:249, 1982). Pollen was collected from at least 100 competitive plants per population, or about 20 plants per parental line, and then mixed with an equal quantity of pollen produced by the standard line W22, characterized by coloured aleurone (dominant marker).
This mixture was then used to pollinate plants of the related F1 (Mu195 x A632). Fifteen and ten well-fertilized ears were considered for each population and line, respectively, and the pollen tube growth rate, relative to that of the standard line, was estimated as the slope of the regression line of the uncoloured kernel frequency on the ear segment number (from 1 [apex] to 5 [base]). Thus, the higher the slope, the faster the pollen tube growth.
The base C2 and C4 showed negative slopes, whose mean was, however, significantly higher than that of Apex cycles (Table 1), indicating a greater growth speed of Base pollen tubes. It should be noted that the slopes of the two Base cycles were slightly higher than that of the better parent (Mu195), whereas the two Apex cycles were intermediate between the two parents.
As regards the sporophytic traits, which were measured on 20 competitive plants per plot, the Base cycles were slightly earlier, with fewer leaves and with shorter plants then Apex cycles. These results indicate that gametophytic selection led to an earliness increase and to a plant size reduction. It is worth noting that the line with faster pollen tube growth (Mu195) was earlier, and with smaller plants, than the other line (A632).
Since an F2 was used as the breeding population, the above correlated changes can be ascribed to pleiotropy (genetic overlap) and/or linkage disequilibrium. To have a clearer picture about the source(s) at work, additional cycles of gametophytic selection are needed.
Table 1. Gametophytic and sporophytic traits of parental lines and selection cycles.
P. Landi and E. Frascaroli
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}