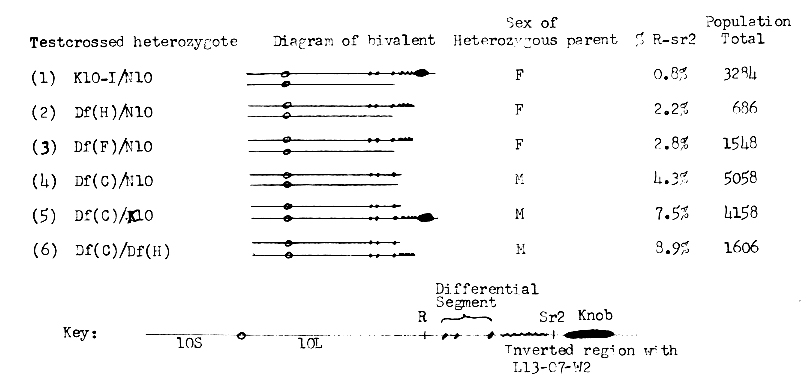
Our studies with abnormal chromosome 10-Type I have shown that the terminal region of the long arm differs from normal 10L in possessing a differential segment of unknown origin marked cytologically by three prominent chromomeres, followed by a euchromatic segment containing the luteus-13, opaque-7, white-2 and striate-2 loci of normal 10L but with the order of W2-07-L13 inverted, and finally a large heterochromatic knob capped by a small euchromatic tip. A number of compounds involving K10-I, N10. and various Df K10-I chromosomes have been constructed in the course of our experiments. Listed below are some of these compounds and the values for R-sr2 recombination detected in testcrosses of each. All of the Df K10-I chromosomes lack a terminal segment of the parent K10-I chromosome including the Sr2 locus. It should be noted that R-sr2 crossing over in K10-I/N10 and all Df K10-I/N10 compounds is restricted to a small homologous region between the R locus and the left end of the differential segment. In compounds involving K10-I/Df K10-I or Df K10-I/Df K10-I, there is a longer homologous segment, and R-sr2 crossovers may occur anywhere between the R locus and the tip of the shorter of the two homologues.
Compounds 1-4 all show a drastic reduction in R-sr2 recombination compared to the value of 35% found in N10/N10 bivalents because of the lack of homology in the distal regions of 10L. The lower crossing over observed in K10-I/N10 heterozygotes as compared with Df(C)/N10 (compounds I vs. 4) might be attributed to the presence of the knob of K10-I in the former compound, since the adverse effect of knob heterozygosity on crossing over in neighboring regions has been well documented in maize. However, a comparison of compounds 2, 3, and 4 shows that a second factor also plays a role in reducing crossing over. All crossovers classified as R-sr2 recombinants occur in the short interval between R and the left end of the differential segment. Recombination in this short interval is highest (4.3%) in Df(C)/N10 compounds and progressively lower (2.7% and 2.2%) in Df(F)/N10 and Df(H)/N10 compounds, owing to the competitive pairing of the inverted W2-07-L13 region with the homologous uninverted region of N10 immediately adjacent to the R locus. Attempted synapsis between the two homologous segments, one of which is both transposed and inverted, will interfere with pairing of the short region distal to R where crossing over occurs. No such interference is found in Df(C)/N10 compounds since Df(C) lacks the critical inverted W2-07-L13 segment. Even in those instances where pairing was accomplished between the standard and inverted W2-07-L13 regions, subsequent crossing over would produce dicentric chromosomes with a low rate of transmission. Thus, Df(F) and Df(H) compounds have a reduced rate of crossing over compared with Df(C), not because of knob heterozygosity since they lack the terminal knob, but because of the presence of the inverted W2-07-L13 segment (or a portion thereof). In K10-I/N10 compounds, both factors are operating to suppress crossing over and an even greater reduction occurs. The effect of the knob alone is apparent in the comparison of K10-I/N10 vs Df(H)/N10 (compounds 1 vs. 2) since K10-I differs from Df(H) mainly by the absence of the terminal knob.
Another example where the knob on K10 appears to play a subordinate role is the comparison of Df(C)/K10 vs Df(C)/Df(H) (compounds 5 vs. 6). In these bivalents the region from R to the tip of Df(C) is homologous and there is no opportunity for competitive pairing since the W2-07-L13 region is present only in the longer homologue and is completely absent from Df(C). In both testcrosses, large populations were scored and in both cases the heterozygous male parent was analyzed. They differ only by the presence of the knob in compound 5. There is 7.5% recombination distal to R in Df(C)/K10-I and 8.9% for the same interval in Df(C)/Df(H) compounds. Little if any reduction due to knob heterozygosity was observed. It is true that the K10 knob is not immediately adjacent or included in the synapsed region where crossing over occurs (it is separated from the synapsed region by the intervening W2-07-L13 segment) and this may prove to be a significant factor.
That crossing over is low per unit length of chromatin in the homozygous differential segment is indicated by our finding of only 8.9% recombination between R and the right terminus of the Df(C) chromosome in Df(C)/Df(H) compounds. Approximately one half of the 8.9% can be ascribed to exchanges between R and the left end of the differential segment, leaving only 4.6% derived from exchanges in the homozygous differential segment. More than 30% of recombination occurs in a region of comparable length in the terminal region of N10/N10 bivalents. Neither the Df(C) nor the Df(H) chromosome has the large knob in the long arm of the K10 chromosome, so it cannot be held responsible for the low incidence of crossing over.
M.M. Rhoades and Ellen Dempsey
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}