A new Adh1 mutant-Adh1-3F1124-was isolated in Berkeley in 1983, by screening F2 families of maize lines that contain Robertson's Mutator (Robertson, Mutat. Res. 51:21, 1978) for anaerobic respiration defects. One family showed segregation of 1/4 of seeds that failed to germinate after 96 hr treatment of anaerobic stress. Seedlings were, however, rescuable by the removal of anaerobic stress immediately following the initial treatment (for detailed screening procedure ref. to C.-H. Chen et al., Maydica, McClintock volume; in press). The inability to grow under anaerobic conditions was attributed to lower expression of ADH enzyme activity in scutellum and root (<5% of its progenitor allele). A homozygous Adh1-3F1124 line was later established by self-pollination of the rescued plants. The most dramatic mutant phenotype of Adh1-3F1124 lies in the observation that ADH expression doesn't seem to be affected in the male gametophyte (pollen grain), although embryonic and somatic tissues continue to show less than 5% of ADH activity. The effect of mutation is therefore organ-specific.
Using DNA probes from the standard Adh1-1S allele, we have constructed a genomic restriction map of the mutant allele. Comparison between the progenitor Adh1-3F allele and the mutant Adh1-3F1124 allele showed that the mutant was characterized by an insertion of about 2 Kb at 5' end of the gene within a PstI and HindIII restriction fragment. This region represents 137 bp 5' to the transcriptional unit, and 204 bp into the Adh1 structural gene. Both the progenitor and the mutant allele have now been cloned into EMBL 3. Preliminary results indicate that the insertional DNA sequence is homologous but not identical to the Mu1 transposable element, which has been previously characterized in other Adh1 mutants isolated from Robertson's Mutator background. We have designated this new element as Mu3. We are now in the process of DNA sequencing and will soon know more about the precise location of the insertion point and the nature of the insertional element. At this point we do not know whether the mutant phenotype is reflected at the transcriptional level, nor do we have any data to speculate on the mechanism of the aberrant organ-specific expression.
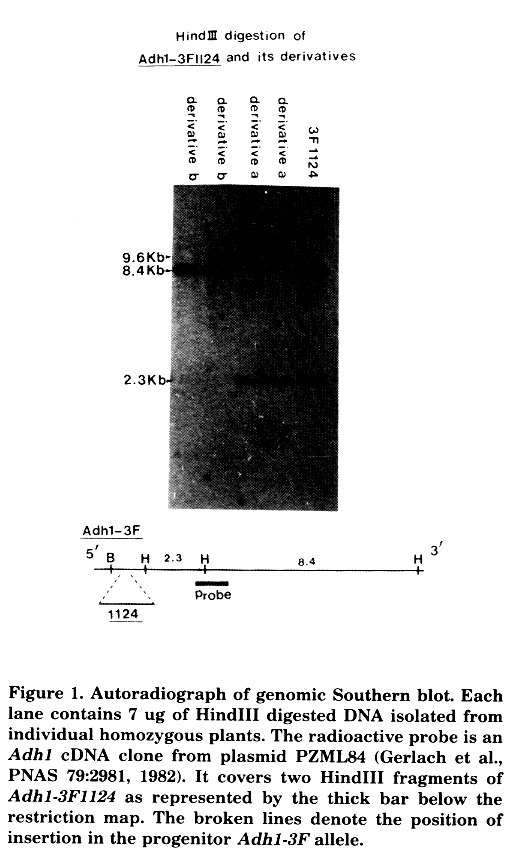
Support of the idea that Adh1-3F1124 is indeed caused by a transposable element insertion is provided by the fact that the allele is genetically unstable both somatically and germinally. Enzymatic staining for ADH activity in the aleurone layer clearly revealed that dark blue single cells or sectors of reverting cells was frequent (on the order of 10-3). Several putative revertant kernels have been isolated (C. Fenczik and B. Kloeckener, unpublished data). Since the original mutant was ADH positive in pollen, ADH null derivatives can be isolated by allyl alcohol treatment, which selects against ADH activity in pollen grains (Freeling and Cheng, Genet. Res. 31:107, 1978). Eleven ADH null derivatives have now been obtained by allyl alcohol selection using a single homozygous Adh1-3F1124 tassel as the male parent. All of them showed stable null phenotype in all the organs and tissues that are known to express ADH. We believe that the derivative null phenotype is due to DNA rearrangement mediated by the Mu3, because all the derivatives show changes in restriction sites within the Adh1 region. Three patterns of rearrangement were detected among 11 derivatives. Figure 1 shows two of them. In derivative-a, rearrangement occurs at the 3' end of the gene where a HindIII site is displaced. This results in an increase of a HindIII fragment from 8.4 Kb to 9.6 Kb. In derivative-b, the 2.3 kb HindIII fragment that comprises 2/3 of the structural gene (intron 1 to exon 8) is missing. This is most likely due to a deletion, since a genomic probe representing from intron 1 to exon 6 of Adh1 also failed to pick up the HindIII-HindIII signal when the same blot was re-hybridized (data not shown). A third kind of rearrangement is now being analyzed.
Figure 1. Autoradiograph of genomic Southern blot. Each lane contains 7 ug of HindIII digested DNA isolated from individual homozygous plants. The radioactive probe is an Adh1 cDNA clone from plasmid PZML84 (Gerlach et al., PNAS 79:2981, 1982). It covers two HindIII frragments of Adh1-3F1124 as represented by the thick bar below the restriction map. The broken lines denote the position of insertion in the progenitor Adh1-3F allele.
Che-Hong Chen, Karen Oishi and Michael Freeling
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}