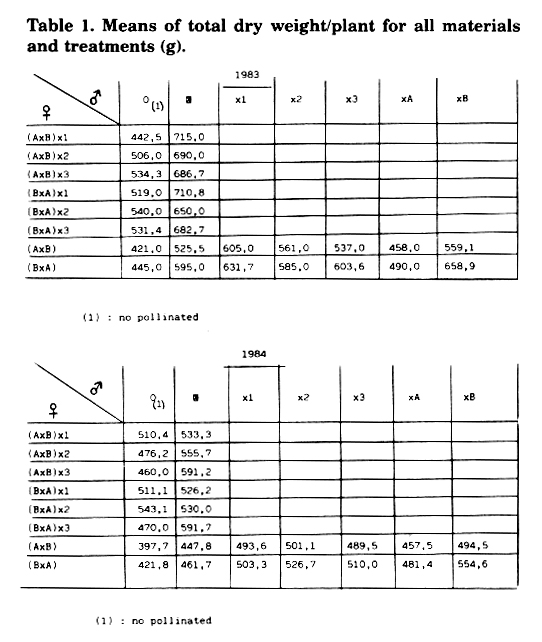
Continuing the work reported in MNL 57: 10-11, we attempted to verify the interaction of environment with manifest events such as reciprocal and xenia effects, and the presence or absence of a similar performance between F1 and F2 progenies in two sets of three-way crosses. The same trial was repeated during two years and for two localities. We used a flint single cross, long cycle opaque-2 (AxB), and the reciprocal (BxA), and two sets of three-way crosses obtained from these single crosses and three dent opaque inbred lines, 1, 2 and 3. The treatments in each single cross were as follows: no pollination, selfing, crossing with 1, 2 and 3 inbreds, and backcrossing with their parental lines, A and B. In both sets of three-way crosses, the treatments were: no pollination and selfing. Experimental design was "randomized plots" of 15 replications. The parameter studied was total dry weight per plant.
In joint analyses measured during 1983 and 1984,
the results (Table 1) were: 1) The reciprocal cross (BxA) is better than
the direct cross (AxB). This result was the same during both summers and
localities. Thus, to analyse all pollination treatments:
| 1983 | 1984 | |
| (AxB) < (BxA) | **P<0.01; D = 9.5% | **P<0.01: D = 5.4% |
2) Only in the three-way cross [(BxA)x3], the total dry weight per plant increased over no pollination with selfing treatment:
1983: no pollination<selfing ***P<0.001; D
= 28.5%
1984: no pollination< selfing *P<0.05; D
= 26.0%
3) In both forms of single cross, the no pollination treatment was always lower than crossing with 1, 2 and 3 inbred lines:
1983: (AxB) no pollination < (AxB)x1,2,3 *P<0.05
(BxA) no pollination < (BxA)x1,2,3 **P<0.01
1984: (AxB) no pollination < (AxB)x1,2.3 *P<0.05
(BxA) no pollination < (BxA)x1,2,3 **P<0.01
4) When the reciprocal single cross (BxA) is pollinated with the parental line B, the total dry weight is better than when the pollination is made with the parental line A. This result is always the same:
(BxA)xB>(BxA)xA *P<0.05
1983 D
= 34.5%; 1984 D
= 15.2%
5) When we compare the F1 and F2 progenies in both sets of three-way crosses, we notice that these hybrids do not have the same performance as far as yield is concerned during two years:
1983: (AxB)x1 > (AxB)x2 > (AxB)x3
[(AxB)x1]F2 > [(AxB)x2]F2 > [(AxB)x3]F2
r = 0.969 *P<0.05
(BxA)x1 > (BxA)x3 > (BxA)x2
[(BxA)x1]F2 > [(BxA)x3]F2 > [(BxA)x2]F2
1984: (AxB)x2 > (AxB)x1 > (AxB)x3
[(AxB)x3]F2 > [(AxB)x2]F2[(AxB)x1]F2
(BxA)x2 > (BxA)x3 > (BxA)x1
[(BxA)x3]F2 > [(BxA)x2]F2 > [(BxA)x1]F2
In summary, we can conclude: 1) The reciprocal effects are invariable, there is not dependence with the environment for these hybrids; 2) The environmental interaction is very important in the xenia effect expression; 3) However, the environmental interaction changes, intensely, the sequence of production in the F1 and F2 progenies.
Table 1. Means of total dry weight/plant for all materials and treatments (g).
R. Fontanet and A. Alvarez
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}