Transposable controlling elements appear to be regular residents of the maize genome, but exist in an inactive form. There is substantial evidence that conditions or agents that induce chromosome breakage promote the activation of transposable elements. These include the genetic induction of chromosome breakage (B. McClintock, CIW Ybk. 49:157-167, 1950; ibid., 50:174-181, 1951; E.B. Doerschug, Theor. Appl. Genet. 43:182-189), X-ray and UV irradiation (M.G. Neuffer, Genetics 53:541-549, 1966; A. Bianchi, F. Salamini, and F. Restaino, MNL 43:91, 1969) and tissue culture (V.M. Peschke, R. Phillips and B.G. Gengenbach, MNL 59:91, 1985). The growing evidence that genetically active elements can be distinguished from inactive ones by their DNA modification patterns (N. Fedoroff, S. Wessler, and M. Shure, Cell 35:235-242, 1983; V. Chandler and V. Walbot, PNAS, in press) raises the possibility that elements are maintained in an inactive or cryptic form by methylation or other modifications of element sequences.
In the present communication, I report some evidence suggesting that Spm elements can themselves promote the genetic activation of previously cryptic elements. McClintock (CIW Ybk. 58:452-456, 1959; ibid., 70:5-17, 1971) showed that an active element could transiently activate an inactive element when present in the same plant, but that the inactive element subsequently segregated from the active one unchanged. The present study was undertaken to investigate the genetic basis of the apparent ability of an altered Spm element, designated a weak Spm (Spm-w) element, to revert to a standard Spm (Spm-s) element. McClintock (CIW Ybk. 56:393-401, 1957; ibid., 62:486-493, 1963) described the isolation of Spm-w elements, which transpose and trans-activate transposition less frequently and later in plant development than is characteristic of the Spm-s element, and reported that such elements can revert to give an Spm-s phenotype (McClintock, Am. Nat. 95:265-277, 1961). I have investigated the genetic basis of this apparent reversion, using an Spm-w derivative of the original a-m5 allele (B. McClintock, CIW Ybk. 60:469-476, 1961).
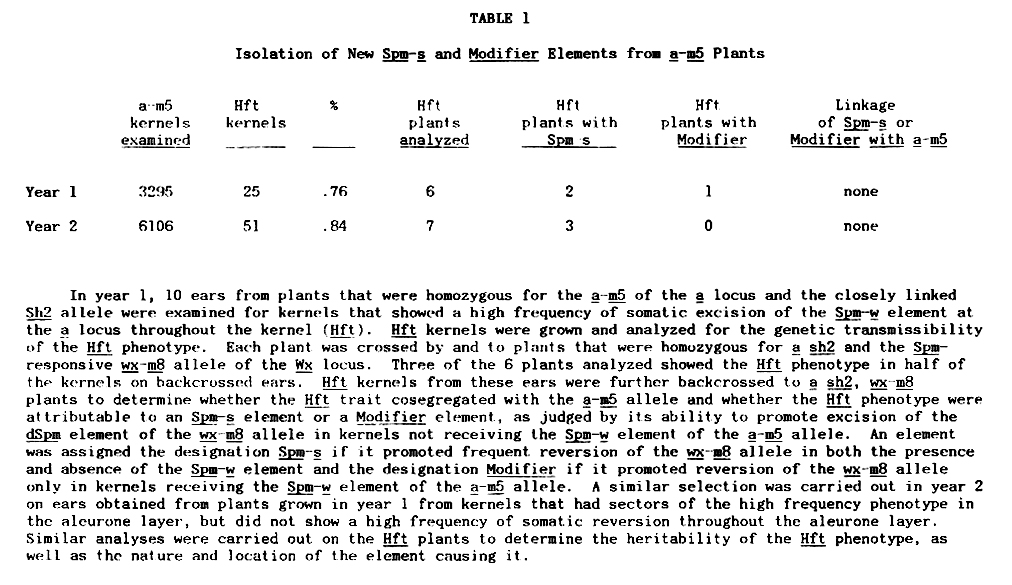
The a-m5 allele used has an Spm-w element inserted at the a locus. The phenotype of the a-m5 allele is colorless with a sparse pattern of small, deeply pigmented A sectors. The Spm-w element at the a locus rarely excises in the germline (<.05%) to give stable null or revertant alleles. However, kernels on plants carrying the a-m5 allele frequently show sectors within which the somatic reversion frequency is much higher than in the remainder of the aleurone layer. Moreover, individual kernels that show a high frequency of somatic reversion (Hft) appear on ears of homozygous a-m5 plants at a relatively high frequency (about 0.8% in the cultures whose analysis is described in Table 1). Plants were grown from a number of Hft kernels and tested for the genetic transmissibility of the Hft trait. Each plant was crossed by and to a sh2, wx-m8 homozygotes. The wx-m8 allele contains a defective Spm element (dSpm) whose excision can be promoted by either an Spm-w or an Spm-s element elsewhere in the genome, but which cannot itself excise from the waxy locus. Almost half of the Hft kernels gave plants in which the Hft trait was genetically transmissible (Table 1). Upon further backcrossing, it was determined that the Hft trait was not linked to the a-m5 allele in any of the plants tested. In 5 of the 6 plants, the Hft trait appeared to be attributable to the presence of an independently segregating Spm-s element, as assessed by its ability to promote somatic reversion of the wx-m8 allele in a sh2 kernels of backcrossed ears. In 1 of the 6 plants the Hft trait showed the genetic behavior characteristic of a Modifier element (McClintock, op. cit., 1957, 1958). Half the kernels receiving the a-m5 allele showed a high frequency of somatic reversion of both the a-m5 and the wx-m8 alleles. None of the a sh2 kernels showed somatic reversion of the wx-m8 allele, indicating that expression of the Hft phenotype was dependent on the simultaneous presence of the Spm-w element of the a-m5 allele.
Thus in all of the plants analysed, the Hft phenotype appears to be attributable to the genetic activation of a previously cryptic element, either an Spm-s or a Modifier. In no case was the Hft phenotype found to be due to the reversion of the Spm-w element at the a locus to an Spm-s element. This is not altogether surprising, since another Spm-w element isolated from the a locus has been found to have an internal deletion within the Spm element (J. Banks, J. Kingsbury, V. Raboy, J. W. Schiefelbein, 0. Nelson, and N. Fedoroff, Cold Spring Harbor Symp. Quant. Biol., in press). The overall frequency of activation of cryptic elements in this experiment is extremely high, approaching 0.4%. Although this type of study has not been carried out yet on other Spm-w alleles, they behave similarly. Several other Spm-w elements in different genetic backgrounds have been examined and show the same tendency to give Hft sectors, as well as Hft kernels at a frequency between 0.1 and 1%.
The spontaneous frequency of activation of cryptic elements can be assessed using an allele such as wx-m8, which has a dSpm element insertion. In a separate study of this type, using a dSpm allele of the a locus, only 2 new genetically transmissible elements were recovered from among more than 100,000 kernels examined. Similar or lower frequencies of spontaneous activation of Dt and Spm elements have been reported by McClintock (op. cit., 1951) and Neuffer (op. cit., 1966). Although controls of this type have not yet been conducted in precisely the genetic background of the present experiments, the frequency with which Hft kernels are detected has been followed in a number of outcrosses and appears to be characteristic of the a-m5 allele, not the genetic background (Table 1). This raises the possibility that the Spm-w element itself promotes the genetic activation of cryptic elements. If this is the case, it appears unlikely that the activation is mediated by an Spm-promoted DNA rearrangement, since the Spm-w element promotes its own excision and that of defective elements at other locations much less frequently (<.05%) than it promotes the activation of cryptic elements (-.4%). A different possibility is that an Spm-encoded gene product can interact with a cryptic element in such a way that it interferes with the maintenance of the cryptic state. A hypothesis that makes use of the recent observations on element methylation is that the cryptic state is maintained by DNA modification and the Spm-encoded gene product can occasionally interfere with modification of newly replicated element DNA, thereby promoting the conversion of a cryptic element to an unmodified (or less modified), genetically active form.
Nina Fedoroff
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}