The mitochondrial DNA (mtDNA) of the S-type cytoplasmically male-sterile maize (cms-S) is characterized by the presence of two linear DNA molecules, S1 and S2, 6.4 and 5.4 kb in length, respectively. In the cms-S versions of most inbred lines examined, these linear molecules are present in nearly equimolar amounts. In four inbred-line backgrounds, however, the cms-S cytoplasm has a ratio of S1:S2 other than 1:1 (J.E. Carlson, Ph.D. thesis, 1983). R825, M825K and M825L cms-S versions all have a fourfold or greater excess of S1 over S2. Cms-S versions of inbred line 38-11, on the other hand, have an average three-fold greater amount of S2 than of S1.
The relative amounts of S1 and S2 in the cms-S cytoplasms have been shown to be under nuclear control (Laughnan et al., Stadler Genet. Symp. 13:93-114,1981). When an inbred-line cms-S version with equimolar amounts of S1 and S2 is recurrently backcrossed as the female parent with M825 as the male parent, the S1:S2 ratio is shifted to the reduced S2 pattern characteristic of M825. This shift does not occur immediately, however, but follows a lag period of one backcross generation; that is, there is no change in the S1:S2 ratio in the F1, a change to an intermediate level of S2 in the first backcross generation, and by the third backcross generation a change in level of S2 equivalent to that of cms-S M825 itself.
The lag phenomenon leads to questions about the number of nuclear genes controlling the relative replication rates of S1 and S2 and about their dominance relationships. Results from Carlson's experiments on genetic control were inconclusive, and new approaches have been taken in attempts to analyze the phenomenon. One of the approaches involves the use of the indeterminate gametophyte gene, ig (J.L. Kermicle, Science 166:1422-24, 1969; Amer. J. Bot. 58:1-7, 1971). The ig mutant increases the rate of occurrence of both maternal and paternal monoploids. The strain of cms-S ig ig used here has the nuclear background of W23 and A158 inbred lines, both of which maintain equimolar amounts of S1 and S2. This stock was crossed as female with M825 and 38-11, and the resulting diploids and monoploids were analyzed for S1:S2 ratios.
Mitochondrial DNA was isolated from young unpollinated ears using a rapid assay procedure developed by Kemble and Bedbrook (Maydica 24:175-180, 1979), and modified by Carlson (Ph.D. thesis, 1983), for use with etiolated shoots.
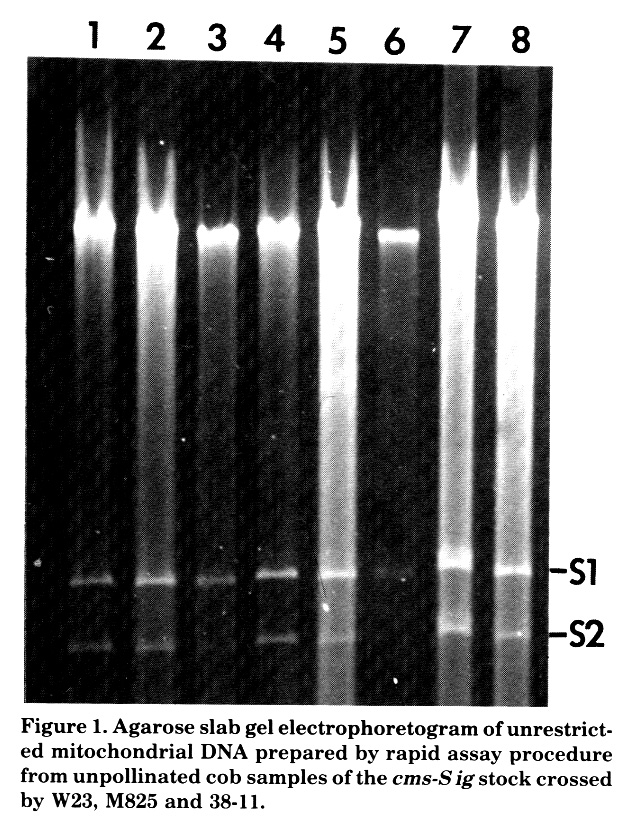
Lane 1 is a mtDNA sample from an RD-WF9/38-11 F1 with a known equimolar ratio of S1:S2; it is included here as a control for comparison with S1:S2 ratios in other lanes.
Lanes 2 and 4 contain mtDNA samples from two W23 maternal monoploids; as expected, the S1:S2 ratio in this nuclear background is approximately 1:1.
Lanes 3 and 6 contain mtDNA samples from S-M825 paternal monoploids. The ratio of S1 to S2 in these monoploids is that characteristic of M825 inbred line, i.e. S1 greater than S2. This is a departure from the previously observed lag in the shift of S1:S2 ratios when cms-S lines with equimolar ratios of S1 to S2 are crossed as female parents with M825. In the case of the M825 paternal monoploids, the nuclear genes of the female parent are replaced by the M825 nuclear genes after only one cross, i.e. in the androgenetic embryo. Recurrent backcrossing of a cms-S equimolar strain by M825 as male parent requires numbers of backcrosses to even approach such a substitution. We discuss the further significance of this below.
Lanes 5 and 8 contain mtDNA samples from S-M825 spontaneously doubled monoploids. The ratios of S1:S2 observed are those characteristic of the M825 inbred line, and again no lag is observed.
Lane 7 contains mtDNA from an S-38-11 paternal monoploid. Cms-S versions of inbred line 38-11 exhibit higher levels of S2 than S1. Contrary to what was observed with the M825 monoploids and the androgenetic diploid M825 derivatives, however, the single 38-11 monoploid does not exhibit a ratio that is characteristic of the 38-11 inbred line; as shown in lane 7, the level of S1 is about the same as S2. Obviously we need to analyze more cases of 38-11 androgenetic derivatives from this cross and follow them through a longer time course.
We had earlier considered that the lag phenomenon might be attributed to a physiological delay associated with the time required for organellar components to adjust to a new equilibrium specified by a change in nuclear genotype, or alternatively, to "recessiveness" of the M825 gene allele(s) associated with the reduced level of S2 replication. The observations on M825 paternal monoploids and on the M825 androgenetic diploids provide no evidence that there is a delay in reaching a newly-established equilibrium; in fact, they strongly suggest that the lag phenomenon observed in the conversion experiments is due to dominance of the nuclear gene(s) controlling normal S2 replication.
Figure 1. Agarose slab gel electrophoretogram of unrestricted mitochondrial DNA prepared by rapid assay procedure from unpollinated cob samples of the cms-S ig stock crossed by W23, M825 and 38-11.
K. K. Kidwell, J. R. Laughnan, S. Gabay-Laughnan
and L.J. Escote
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}