S-type male-sterile cytoplasm in maize is characterized by the presence in the mitochondrial genome of two autonomously replicating linear plasmid-like DNA species called S1 and S2, 6.4 kb and 5.4 kb in length, respectively. In the S-male sterile M825 inbred line background, cytoplasmic reversion to fertility is accompanied by the loss of these plasmid-like DNAs and by rearrangements in the main mitochondrial genome that involve sequences homologous to S1 and S2 (Levings et al., Science 209:1021-1023, 1980; Laughnan et al., Stadler Genet. Symp. 13:93-114, 1981; Kemble and Mans, J. Molec. Appl. Genet. 2:161-171, 1983; Schardl et al., Cell 43:361-368, 1985). In the WF9 inbred line background cytoplasmic reversion to fertility does not involve the loss of the free S1 and S2 plasmid-like DNAs (Escote et al., MGCNL 59:100, 1985; Plasmid, in press; Ishige et al., MGCNL 59:98-99, 1985; Japan J. Breed. 35:285-291, 1985), and the pattern of rearrangement of sequences homologous to S1 and S2 in the main mitochondrial genome is distinct from that of M825 revertants. Revertants that arise following conversion of the RD- or ML- WF9 inbred-line background to the M825 nuclear background by recurrent backcrossing lose the free S1 and S2 episomes, and exhibit a rearrangement of the main mitochondrial genome that is characteristic of M825 revertants rather than of WF9 revertants (Escote et al., MGCNL 59: 100, 1985, and unpublished results). This indicates there is a major effect of the nuclear background on the organization of the mitochondrial genome of the fertile revertants. It was not clear, however, whether the nucleus exerts this influence on the mitochondrial genome organization upon reversion or prior to the reversion event. We therefore studied the organization of the mtDNA of the progeny in each generation of the conversion of the male-sterile RD-WF9 to the M825 nuclear background using as mtDNA probes the cloned S1 and S2 sequences, and the cloned mitochondrial genes for cytochrome oxidase subunits I (COI) and II (COII) and apocytochrome B (COB).
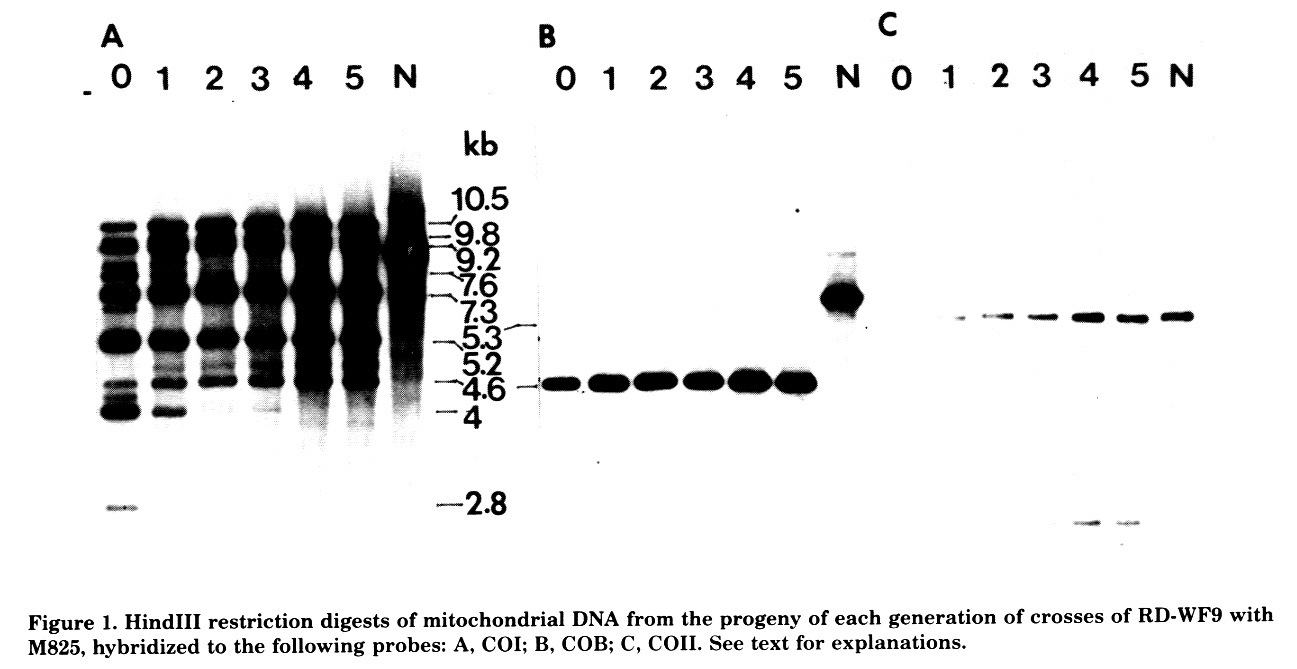
Figure 1 shows HindIII digests of mitochondrial DNA from the progeny of five generations of backcrosses of RD-WF9 by M825, hybridized to the probes COI (Panel A), COB (Panel B), and COII (Panel C). Lane 0 corresponds to the female parent RD-WF9, lanes 1 to 5 correspond to the progeny of the first to the fifth cross of RD-WF9 by M825, and lane N to the normal fertile maize mitochondrial genome. As shown in Figure 1, reorganization involving the COI gene is apparent as early as the F1 generation, with the loss of three COI fragments originally present in the RD-WF9 parent (refer to figure for approximate sizes of the fragments). In the progeny of the first backcross to M825, two more COI bands disappear. Accompanying the loss of these COI bands is the gradual development of a COI band of about 4.8 kb that is not detectable in the RD-WF9 female parent. The pattern of COI gene organization observed in the progeny of the first backcross by M825 persists through the progeny of the fourth backcross, except for the increasing level of the 4.8 kb band in later generations. A similar pattern of change is observed with S1 and S2 probes, although in this case the change is limited to early loss of bands without the accompanying appearance of a new band (data not shown). With the COB and COII probes (Panels B and C, respectively) there is no apparent alteration in organization accompanying the conversion process.
These results indicate that the influence of the M825 nuclear genome on mtDNA organization can occur in the absence of reversion. The changes appear to involve limited areas of the main mitochondrial genome, i.e. S1 and S2 regions of integration, and COI gene sequences but not COIl and COB genes. The changes involve the loss of bands originally present in the RD-WF9 female parent, and the appearance of a new band that was not apparent in the original RD-WF9 parent. The pattern of changes observed is compatible with the model of a multipartite mitochondrial genome (Lonsdale et al., Nucl. Acids Res. 12:9249-9261, 1984) composed of different replicons with varying contents and sequences, e.g. those of S1, S2 and COI. Given that the WF9 nuclear genome is characterized by a specific distribution and content of such variant replicons, the M825 nuclear genotype appears to establish a different equilibrium that favors the replication of some of these elements, suppresses that of others and leaves still others unchanged.
Figure 1. HindIII restriction digests of mitochondrial DNA from the progeny of each generation of crosses of RD-WF9 with M825, hybridized to the following probes: A, COI; B, COB; C, COII. See text for explanations.
L. J. Escote, S. Gabay-Laughnan and J. R. Laughnan
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}