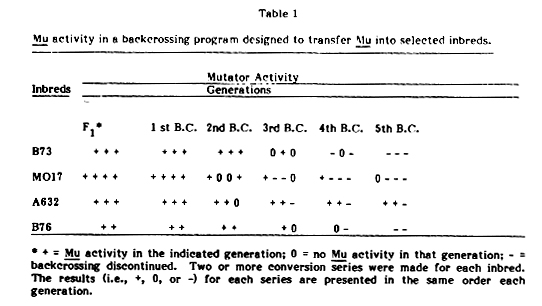
Early in our research program we desired to transfer the Mu system into the inbred lines B73, Mo17, A632 and B76. Each generation of backcrossing was tested for the retention of an active Mu system. However, only very rarely was it possible to retain mutator activity in these backcrossing programs after a few backcross generations, in spite of selecting each generation for crosses with the most active Mu parents. Table 1 lists the inbreds tested and the number of backcross generations before Mu activity was lost. The generation of loss varied somewhat from inbred to inbred, but in all inbreds, except for two series of crosses with A632, Mu activity was gone by the fifth backcross generation.
In the early days of my work with the y9 stock, which was the original source of Mu, I had transferred the y9 gene, by a series of backcrosses, into the inbreds W22 and M14. When I began my first systematic investigation of the mutator system, I tested most of my y9 stocks from different backgrounds for this activity, including the W22 and M14 y9 stocks. These latter two stocks did not have mutator activity, while most other stocks of y9, which had been maintained in more or less heterogeneous backgrounds, showed the mutator phenotype. Thus, the M14 and W22 conversions probably represent two additional instances where mutator activity cannot be sustained in an inbred background.
These results may have significance in light of recent observations in laboratories at Stanford, Purdue and Iowa State University that some inbred plants have modified Mu1-like sequences and, hence, presumably possess the system for modifying the Mu1-like elements. Such active Mu-modifying systems might be expected to inactivate the Mu1-like elements contributed by the Mu parent in my attempts to transfer this system to inbred lines.
In 1983, my working hypothesis was that the loss of Mu activity was due to the inability of the "inbred condition" to support the transposition of Mu elements. To test this hypothesis, plants from the first Mu-Ioss backcross generation were crossed with a different inbred to produce a vigorous F1 (e.g., B73 Mu-Ioss x Mo17 etc.). Selfed ears of plants from these F1 progenies were scored for new mutants that would be observed if either parent had Mu activity, but no new mutants were found. These results are as expected since neither parent has an active Mu system. In 1984, these F1's were outcrossed to an unrelated standard line. If restoring vigor to these inbred-Mu-Ioss backcross stocks activates the Mu system, mutants should be observed in the progenies of the selfs. Other phenomena might also activate the Mu system, such as a hybrid dysgenesis-like phenomenon in these F1's, or possibly deactivation of the modification systems for either or both parents. These selfs were made last summer, but the ears are yet to be classified and the seedling tests made.
The seeds from these outcross plants were sown this summer and the plants selfed. If the Mu system has been reactivated, some of these self progeny should segregate for new mutants.
Table 1. Mu activity in a backcrossing program designed to transfer Mu into selected inbreds.
Donald S. Robertson
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}