In the literature, Mu activity is measured by two criteria. The standard measure of this activity is in the germ line as indicated by the induction of mutants. More recently, with the advent of unstable (mutable) Mu-induced aleurone mutants, activity is often measured by the frequency of spots. These two phenomena may or may not be measuring the same thing. There is good evidence that events associated with germinal activity involve replicative transposition, while somatic instability is probably the result of excision events. The possibility that excision events are sometimes involved in germinal activity has not been ruled out. The following experiments bear on the relationship between germinal and somatic activity of Mu.
Last year (MGCNL 59:11-13, 1985) we reported on the possibility of restoring somatic mutability to stable mutant derivatives of Mu-induced a1 mutants. This was done by crossing stable al derivatives with active purple-aleurone-Mu (Pl Mu) stocks and then selfing the F1. In many of these selfed progeny, the a1 seeds had regained mutability. In several of the crosses, the Pl Mu parent had lost Mu activity for the first time (i.e., had become a first time Mu-loss stock). In some crosses to a stable al derivative, where such Pl Mu-loss stocks were used, the selfed F1 ears segregated for mutable seed. The a1 stable lines were also outcrossed as males to non-Mu purple aleurone stocks. The selfed ears from these outcrosses segregated for predominantly stable seeds. This demonstrated that the stable condition was inherited. These latter self progeny were seedling tested to determine if new mutants were being induced in the stable parents. In none of these tests were new mutants observed. Thus the loss of somatic instability seems to be associated with the loss of germinal activity. Is the reverse true? The cross mentioned above, in which somatic mutability was restored to a stable a1 derivative by crossing to a Pl Mu stock that had lost germinal mutator activity, will provide information bearing on this question. In this situation, both the a1 stable derivative line and the Pl Mu line have been shown to lack germinal activity. Thus, neither parent had a functional germinal Mu system.
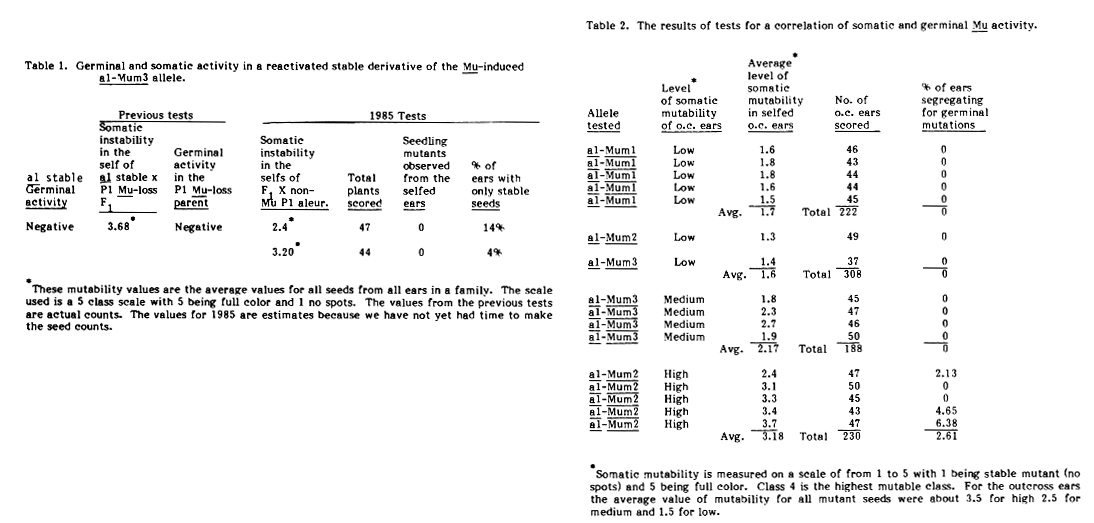
However, the F1 has recovered somatic instability. If the two phenomena (i.e., somatic and germinal instability) are related or linked in some functional way, the F1 plants might be expected to recover germinal Mu activity as well as somatic mutability This was tested by outcrossing the a1-Mum3 stable /Pl Mu-Ioss F1 ( in which somatic instability is recovered) to a non-Mu purple line and selfing the outcross ears, scoring the segregating a1 seeds for somatic mutability again and seedling testing the self progeny to determine if new mutants, expected in the progeny of a plant with a germinally active Mu system, are occurring. The results of such tests, along with the earlier results of the tests of the F1 and parents, are given in Table 1. These admittedly limited data suggest that not only was somatic instability restored to the stable allele, but the mutable condition is now inherited. Only an occasional ear is seen in the outcrosses that does not segregate for mutable seeds. More importantly, the restoration of Mu activity in the aleurone does not appear to be accompanied by the restoration of germinal instability.
A second approach for answering the question about the relationship between somatic and germinal activity was to determine if there is a correlation between the extent of somatic instability, as measured by the frequency of spotting in aleurone mutants, and germinal activity, as measured by the induction of mutations. Selfed ears of outcross progenies of a1-Mum1, a1-Mum2 and a1-Mum3 with low, medium and high levels of somatic activity were scored for the level of somatic mutability and germinal mutator activity as measured by the frequency of the selfed ears segregating for new mutants. The outcrosses were to a1 sh2. Results are shown in Table 2. Some, but not all, of the parents of the plants that gave high average somatic mutability also had germinal Mu activity. None of the parents giving rise to low or medium outcross ears had germinal Mu activity. Thus, the mere presence of a system capable of causing somatic mutability is not sufficient for germinal activity of the Mu system. The two activities of the Mu system do not seem to be completely unrelated, however, since the greater the somatic activity the greater the likelihood of having germinal activity. The observation that stocks of stable derivatives of mutable Mu-induced mutants do not have germinal activity also suggests a relationship with respect to the loss of germinal mutations from stable derivative lines. It should be noted, however, that this is not the case with stable Mu-induced mutants that are stable when first observed. They usually have germinal Mu activity. Thus the loss of germinal activity in stable derivatives of mutable alleles is not the result of the stable phenotype per se.
When one considers what is known about factors affecting germinal Mu activity and somatic activity there are some obvious differences. Bennetzen (In: Plant Genetics vol. 35, in press) has shown that there is little correlation between Mu1 copy number and the somatic reversion frequency (level of somatic mutability). In somatic mutability the level of activity ultimately depends upon what happens to the Mu component at the locus (i.e., does it excise or not and how often, etc.). However, in germline mutations the level of activity depends heavily on how many loci receive a transposed element. Thus, the more elements that can move, the greater the number of gametes with mutants. Therefore in somatic events we are looking at the factors influencing the movement of only one element, while in the classical germline events many transpositions are involved. The observations that somatic mutability can be turned on again while leaving germline mutability off (Table 1) and that the level of somatic mutability is not closely correlated with germinal activity (Table 2) suggest there are some differences in the two phenomena. The observation that the loss of somatic mutability is linked with the loss of germinal activity, on the other hand, suggests some similarities.
Table 1. Germinal and somatic activity in a reactivated stable derivative of the Mu-induced a1-Mum3 allele.
Table 2. The results of tests for a correlation of somatic and germinal Mu activity.
Donald S. Robertson
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}