We recently listed the isozyme loci being studied by starch gel electrophoresis in our laboratory and summarized the available mapping data in a series of reports (Wendel et al., MGCNL 59:87-90). Subsequent work has resulted in further clarification of the chromosomal locations of markers on chromosomes 1 (Acp4, Dia2), 6 (Adk1), 8 (Tpi3), and 10 (Sad1). Previously unreported information on these loci and their chromosomal locations follows:
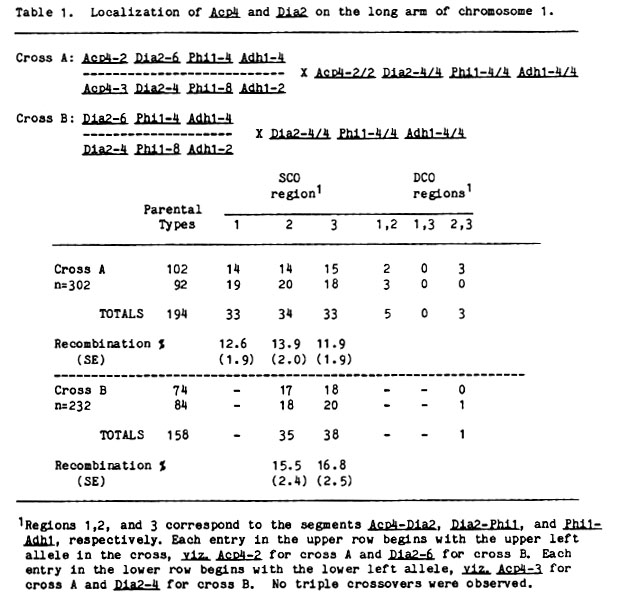
Chromosome 1 - Seven isozyme loci have been mapped on the long arm of chromosome 1: Amp1, Mdh4, mmm, Pgm1, Adh1, Phi1, and Gdh1. Joint segregation of several of these loci with two new isozyme markers, Dia2 (Wendel et al., MGCNL 59:87-88) and Acp4 (Kahler, J. Hered. 74:239-246), suggested that these loci also reside on chromosome 1. Their placement on 1L was confirmed by the testcross data of Table 1. These data indicate that Acp4 and Dia2 are distal to all previously mapped isozyme loci on 1L, and collectively lead to the following map for the four most distal isozyme markers:
Acp4 - 12.6 - Dia2 - 14.6 - Phi1 - 14.0 - Adh1
Phi1 is currently thought to reside approximately at map position 140, and consequently, Dia2 and Acp4 should map to positions 155 and 167, respectively. Acp4 may thus be the most distal marker known on 1L. Crossing experiments between Acp4, Ts6 (tassel seed, 1-158), and bm2 (brown midrib, 1-161) are underway in order to clarify the gene order among these terminal markers.
Chromosome 6 - Some preliminary data suggested that Adk1, the gene encoding adenylate kinase isozymes, segregated non-randomly with other isozyme markers on chromosome 6. Testcrosses of plants heterozygous for various combinations of the loci Adk1, Pgd1 (phosphogluconate dehydrogenase), Enp1 (endopeptidase), and Hex2 (hexokinase) confirm that Adk1 is on chromosome 6 (Table 2). Averaged across families (Kramer and Burnham, 1947; Genetics 32:379-390), there is 14.8% ± 1.9% recombination between Adk1 and Pgd1. These and previous data suggest the following gene order and map distances for isozyme markers on 6:
Adk1 - 14.8 - Pgd1 - 3 - Enp1 - 42 - Hex2 - 43 - Idh2 - 2 - Mdh2
The gene order among Adk1, Pgd1, and Enp1 is consistent with the data of Table 2, and is additionally supported by previous work (Wendel et al., MGCNL 59:89-90). Because the B-A translocation stock TB6Lc uncovers all markers distal to Pgd1, and because the TB-6Lc breakpoint is somewhere in the vicinity of the centromere (perhaps between 6-4 and 6-9) the above data suggest that Adk1 is near the centromere, and perhaps even on 6S. It should be noted, however, that the recombination estimates between Adk1 and Pgd1 are heterogeneous across families, and consequently the distance between these two genes and the location on 6S for Adk1 must be considered tentative. We are currently attempting to map all isozyme markers on chromosome 6 with respect to the morphological markers rgd1 (ragged seedling, 6-4), Y (yellow endosperm, 6-13), pg11 (pale green, 6-34), su2 (sugary, 6-54), and py1 (pygmy plant, 6-65).
Chromosome 8: Previous work has demonstrated that there is approximately 23% recombination between Idh1 (isocitrate dehydrogenase) and Mdh1 (malate dehydrogenase) on chromosome 8 (Goodman et al., Genetics 96:697-710). We crossed the B-A translocation stock TB-8Lc as male onto an Idh1 -Mdh1 tester and uncovered Idh1 (hypoploids recovered in 4 of 50 plants examined) but not Mdh1, suggesting that Idh1 is on 8L, and Mdh1 is either on 8L near the centromere or on 8S. Analysis of an F2 segregating for Idh1, Mdh1, and Tpi3 (triose phos'phate isomerase) confirms the previous estimate of recombination between Idh1 and Mdh1 (30.0% ± 2.6%, and additionally suggests that Tpi3 is on 8S approximately 22 map units from Mdh1 (Table 3).
Tpi3 thus becomes only the second factor (along with Bif1, barren inflorescence) mapped on the short arm of the most poorly marked chromosome of maize. With the available data it is not possible to determine whether Mdh1 is on 8S or 8L. Testcrosses of the three isozyme loci with the morphological markers fl (floury, 8-0) and j1 (japonica striping, 8-42) will be analyzed in 1986.
Chromosome 10: An additional isozyme marker on chromosome 10 was confirmed through the analysis of the selfed progeny of an F1 that was heterozygous for both Glu1 (P-glucosidase) and Sad1 (shikimate dehydrogenase). These data (Table 3) demonstrate tight linkage between Glu1 and Sad1, with 1.0% ± 0.5% recombination. Efforts are being made to map these markers relative to various morphological markers on chromosome 10.
Table 3. Joint F2 segregation data and maximum likelihood estimates of the recombination fractions (r) for three isozyme loci on chromosome 8 and for two isozyme loci on chromosome 10.
Jonathan F. Wendel, Major M. Goodman and C.W. Stuber
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}