Plant phenotype is closely associated with endosperm protein content in the progeny of hybrids between perennial teosinte and maize. Maizoid plants have lower protein content than those of intermediate phenotype, and the latter have a lower level than teosintoid phenotype (see MNL 59:69, 1985).
With the aim of establishing the existence of other ways of associations between plant morphological phenotype and kernel storage proteins, a more detailed study was done of F2 and F3 samples derived from perennial teosinte (Z. perennis) crossed with Gaspe. Twelve plants were chosen: 1 to 4 with teosintoid phenotype, 5 to 8 with intermediate phenotype and 9 to 12 with maizoid phenotype. Endosperm was separated from kernels of each one of these plants, and was used to fraction storage proteins in accordance with their solubility (Landry-Moureaux, 1970). Alcohol-soluble fraction (zein) was used to separate molecular components through polyacrylamide gel electrophoresis, following the Sastry and Virupasksha (1967) technique. Results obtained are shown in Table 1 and Figure 1. Protein pattern variability between plants is considerable. Not only are parental patterns partially recovered but also intermediate protein patterns and transgressive ones are produced (see Table 1). This fact clearly points out the enormous variability engendered as a product of germplasm recombination of distant species. At the storage protein level, this variability corresponds with that morphological variability that has been largely stated in previous articles published in this Newsletter. Zein electrophoretic patterns do not show such a high variability (see Figure 1). This fact may be due, perhaps, to the absence of a great differentiation between parental species for genes that code for zein (MNL 55:62) and the main differentiation may reflect the action of regulatory genes more than structural genes.
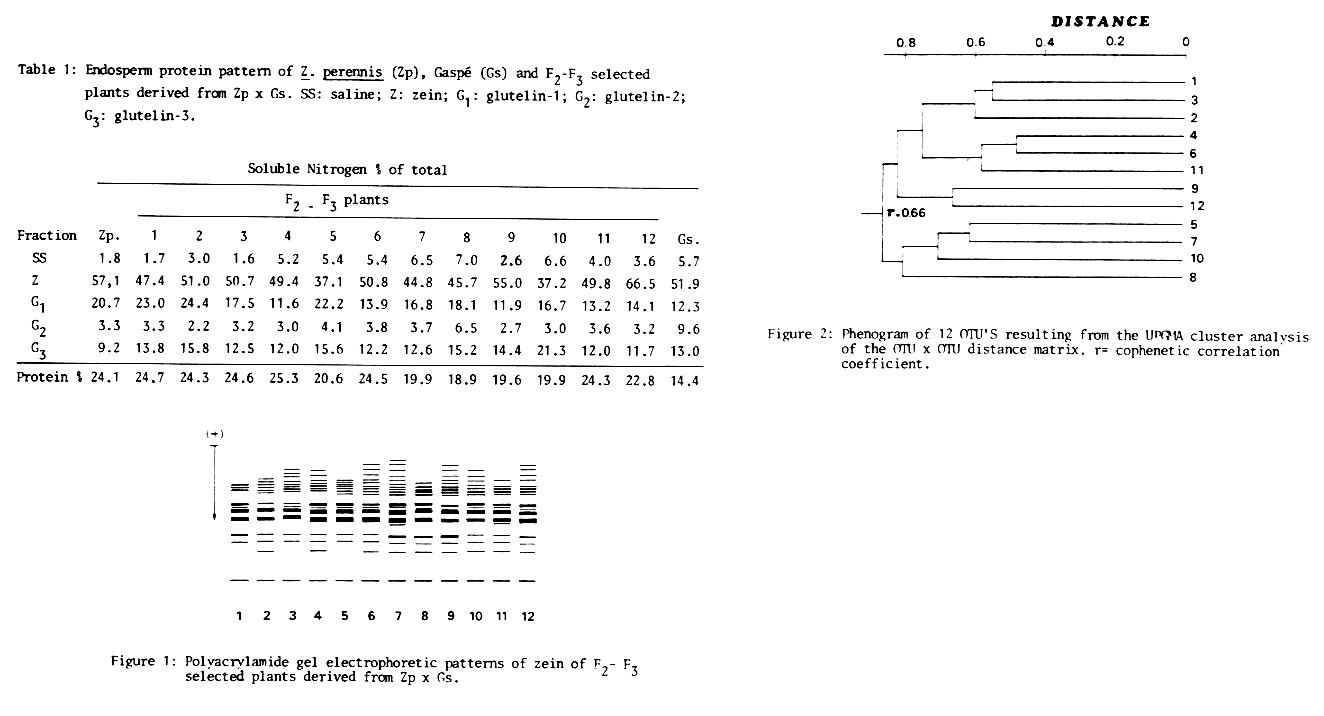
A basic data matrix (BDM) was built with the data present in Table 1 and Figure 1, and was used to calculate the distance matrix among the 12 individuals studied, which constitute the 12 OTU's. The phenogram shown in Figure 2 was derived by "Mean Taxonomic Distance" applying the UPGMA method.
In some cases storage proteins keep a tight relation with the plant morphological phenotype. This occurs in the cluster formed by 1, 2 and 3 teosintoid plants. Plants 4, 6 and 11 belong to different morphological groups, denoting that a partial character recombination has taken place. Plants 9 and 12 are maizoid and are linked between them by their relation to storage proteins. In the 5, 7, 10 and 8 cluster there are 3 plants with intermediate morphological phenotype and one (10) maizoid, pointing to recombination.
The results obtained show in some cases the existence of a height association between plant clustering by morphological traits and by storage proteins. Otherwise no congruence exists in clustering. Therefore it may be inferred that specific traits, both morphological and biochemical, which differentiate perennial teosinte from maize, are mainly inherited in an independent way. Bearing in mind that perennial teosinte and maize are the basic species of genus Zea (see MNL 59:61, 1985) results presented constitute another evidence that these species carry all the genetic information necessary to produce the main portion of possible variants in storage proteins, and that are in some instances the patterns of the remaining taxa.
Liliana Ferrari and Jorge L. Magoja
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}