Most variation in maize has been produced or maintained by teosinte introgression. There is considerable documentation about the important role that teosinte has played in the evolution of modern maize. The idea of employing maize wild relatives to improve the cultivated species is not novel either in its theoretical conception or practical implementation. However, in view of the interesting results obtained from our perennial teosinte (Z. perennis) germplasm introgression into the maize project, it seems convenient to report the experience reaped since the beginning of the research in 1983.
Perennial teosinte (Z. perennis) and diploperennial teosinte (Z. diploperennis) have been our donors of wild germplasm. These species were chosen because they are the most primitive taxa of the genus and thus are the ones that produce more variability and heterosis in crosses with maize. Our main objective was to produce a controlled introgression of teosinte into maize, so as to transfer into the cultivated species those genes or alleles able to produce additional heterosis and increase genetic variability. Now we report results of perennial teosinte germplasm introgression. This is the project that raises more expectancy and where initially the main difficulties of practical implementation have been manifested.
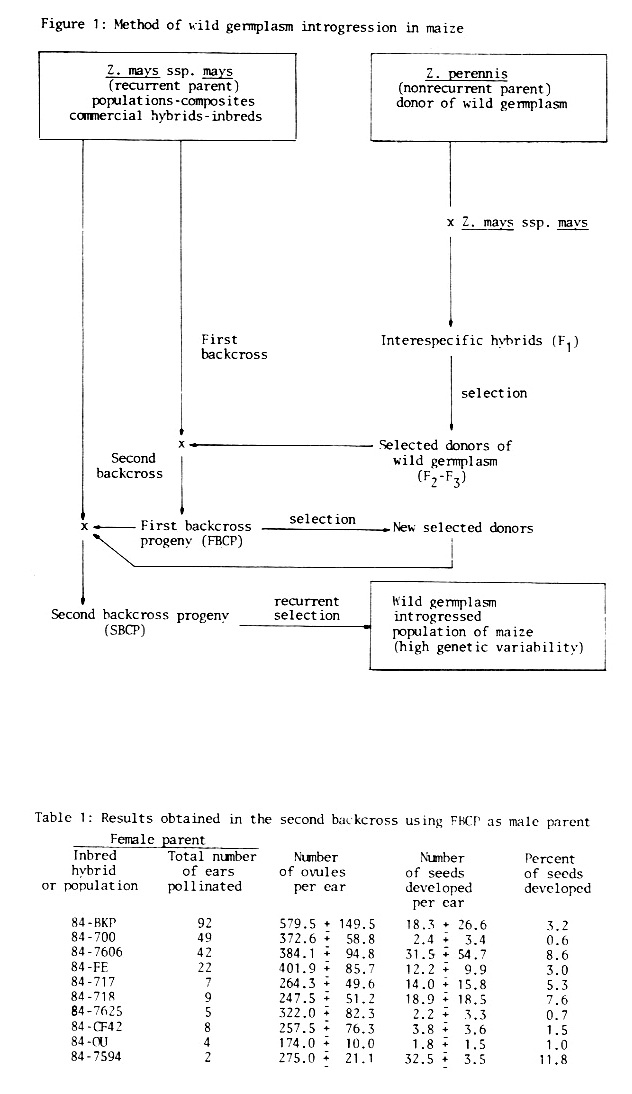
As is shown in Figure 1, Z. perennis was the wild germplasm donor and maize was the recurrent parent.
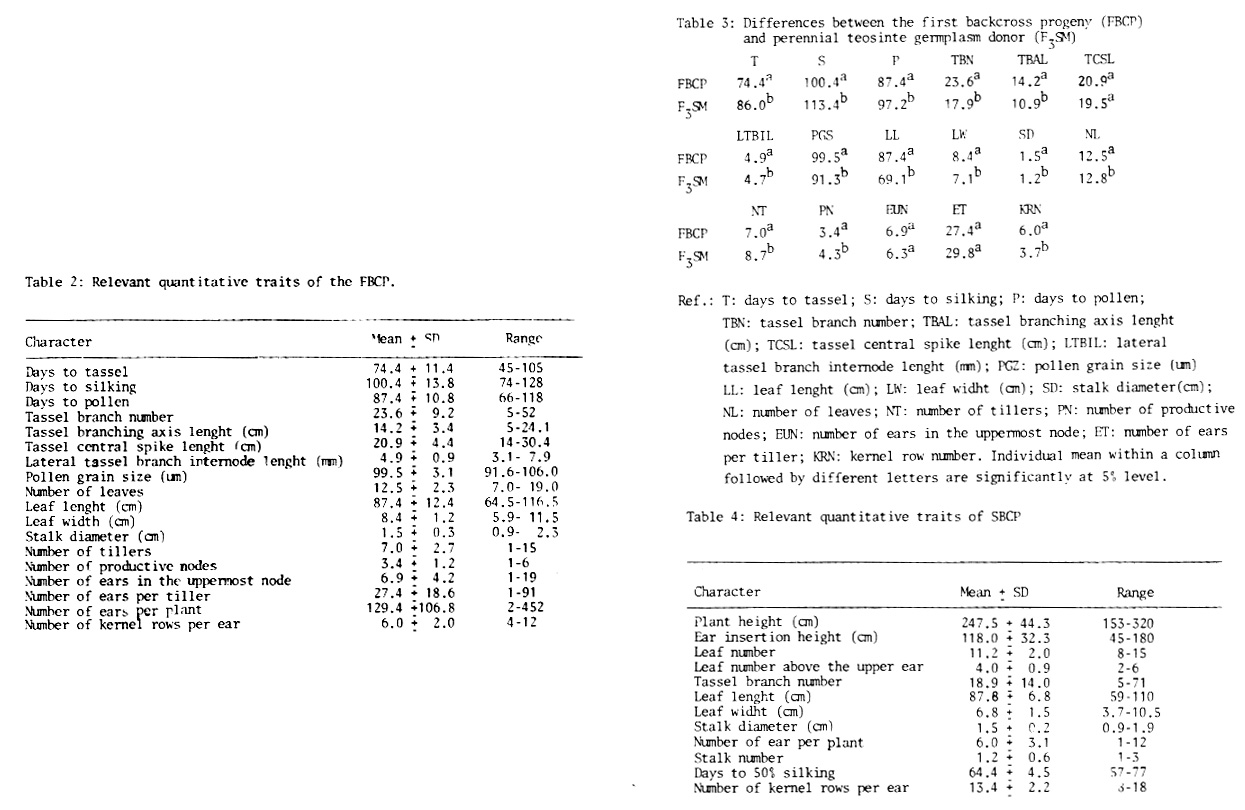
Since hybrids between perennial teosinte and maize have low fertility and produce few viable kernels when crossed again with maize, the donating material was previously selected. So, individuals with maizoid phenotype, highly vigorous and fertile, were chosen among F2 and F3 progenies and used in the first backcross. Seed development was low (15% of available ovules) when maize ears were pollinated with pollen from perennial teosinte germplasm donating individuals, and only 10% of them or less were viable. However, the plants obtained were highly heterotic and showed a high genetic variability The next year, a great number of maize ears were pollinated with a mixture of pollen collected from several plants of the first backcross progeny (FBCP). Maize pollen was also used to pollinate FBCP plants.
We thought that after the first backcross more seeds should be obtained, but surprisingly only 4 kernels for each 100 available maize ovules were obtained from the new pollinations (see Table 1). However, our surprise was even greater when, cultivating the second backcross progeny (SBCP), almost every seed was viable and produced very vigorous plants which presented a high variability within them. Since all maize traits were mainly recovered in the plants obtained after the second backcross, we have considered it appropriate to interrupt backcrosses and start a recurrent selection process with the aim of forming a maize population with wild germplasm introgression that may be used in the future to obtain inbred lines. During the application of this method we have evaluated 30 traits of agronomic and/or specific importance in the donating materials as in the progenies of the backcrosses obtained. These characters have been used to point out the progress in recovering the specific maize traits and to quantify the variability produced by wild germplasm contribution. We shall give only a few results obtained with the aim of showing that changes operated during the introgression process and variability occurred in many of the evaluated characters.
Table 2 shows the results obtained for some characters evaluated in FBCP. It is important to remark on their high variability. In general FBCP plants have recovered enough maize specific traits, but they also keep undesirable traits such as numerous small ears per productive node and many stalks per plant. In general FBCP plants are very vigorous and extremely prolific, although ears produced are generally small and badly covered by husks. In spite of the fact that desired plant traits (that is to say plants like maize but with high vigour and prolificity) have not been reached yet, the progress towards our objective was very significant. This may be proved with the results shown in Table 3, where several FBCP traits are compared with perennial teosinte germplasm donating material (F3SM).
At this moment SBCP is being cultivated in the field. All the same, we have some preliminary results available from this SBCP cultivated in the greenhouse (see Table 4). Although the evaluated traits are few and come from a small SBCP population, they are useful to demonstrate the extremely high engendered variability. The most important fact is that SBCP plants have recovered all maize traits, preserving in most cases an exceptional heterosis expressed through their prolificity. Although the number of ears per plant is lower than in FBCP, they are bigger and well covered by husks over vigorous and strong stalked plants. The variability engendered is enormous: from very poorly developed plants with thin stalks and few small ears to strong stalked plants with 2 to 5 ears, which are bigger and well formed and add as desirable traits simultaneity in pollen shed and silk emergence, and earliness given from Gaspe germplasm (parent of original crosses). SBCP results cannot be compared with FBCP ones, especially in plant size, stalk diameter, etc., because greenhouse grown plants are higher and have thinner stalks and smaller leaves. Nevertheless the results obtained show a real recovery of maize traits after the second backcross. Simultaneously, high diversity that will allow us to obtain in the future improved maizes from materials with perennial teosinte introgression have been produced. Most outstanding SBCP individuals have high yield power and therefore an appropriate selection of them may give a significant increase in kernel yield of current commercial maize.
Jorge Luis Magoja and Gabriela Pischedda1
1Estac. Exper. Reg.
Agrop. Pergarnino-INTA- Pergarnino, Bs, As., Argentina
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}