When the evolutionary study of Zea began in the last Newsletter (MNL 59:61, 1985) we stated that clusters based only on fruit traits were not congruent with those based on other traits. We also stated that this might be due to the inclusion of some non-specific fruit traits. The aim of this article is to present a reexamination of the clusters based on fruit traits. So, we have used basically the same data previously employed but have included some other data. Our objective is to determine an appropriate selection of fruit traits, morphological and biochemical, that may lead to clustering congruent with clusters based on other plant characters. The same 7 operational taxonomic units (OTU's) were studied. Sixteen traits were scored for each OTU as follows: (1) Kernel (PG) enclosed (1), naked (0); (2) Kernel (TG) small (0), intermediate (1), large (2): (3) Cupulate fruit case (FCFC) trapezoidal (2), triangular (1), horizontally compressed (0); (4) Cupule orientation (OC) vertical (0), horizontal (1); (5) Kernels per cupule (GC) one (1), two (2); (6) Number of fruit cases per spike (NCF); (7) Endosperm protein content (%) (PE); (8) Landry-Moureaux saline soluble proteins (PSS); (9) Glutelin-1 (Gl); (10) Glutelin-3 (G3); (11) Pericarp thickness (µm) (EP); (12) Aleurone layer thickness (µm) (ECA); (13) Starch granule size (µm) (TGA); (14) Zein body size of zone I (µm) (TGZI); (15) Zein body size of zone 2 (µm) (TGZ2); (16) Zein body size of zone 3 (µm) (TGZ3). Given a basis data matrix (BDM) (Table 1) of 16 characters by 7 OTU's the data were analyzed by cluster analysis. The BDM was standardized (BDMS) by characters to remove the unequal weights imposed on it by the use of different scales of measurement. Phenograms were derived by (1) the Pearson product-moment correlation coefficient, applying the unweighted pair group method using arithmetic averages (UPGMA) (Figure 1), (2) the "Mean Taxonomic Distance" between pairs of OTU's served as input in the calculation by UPGMA (Figure 2) and by the COMPLETE cluster analysis (Figure 3). A character x character correlation matrix was obtained from BDMS by calculating the Pearson product moment correlation coefficient between each pair of the 16 characters, and served as input in the calculation of a phenogram by UPGMA (Figure 4). The "Manhattan Distance" between pairs of OTU's served as input in the calculation of "Wagner Trees" (Figure 5).
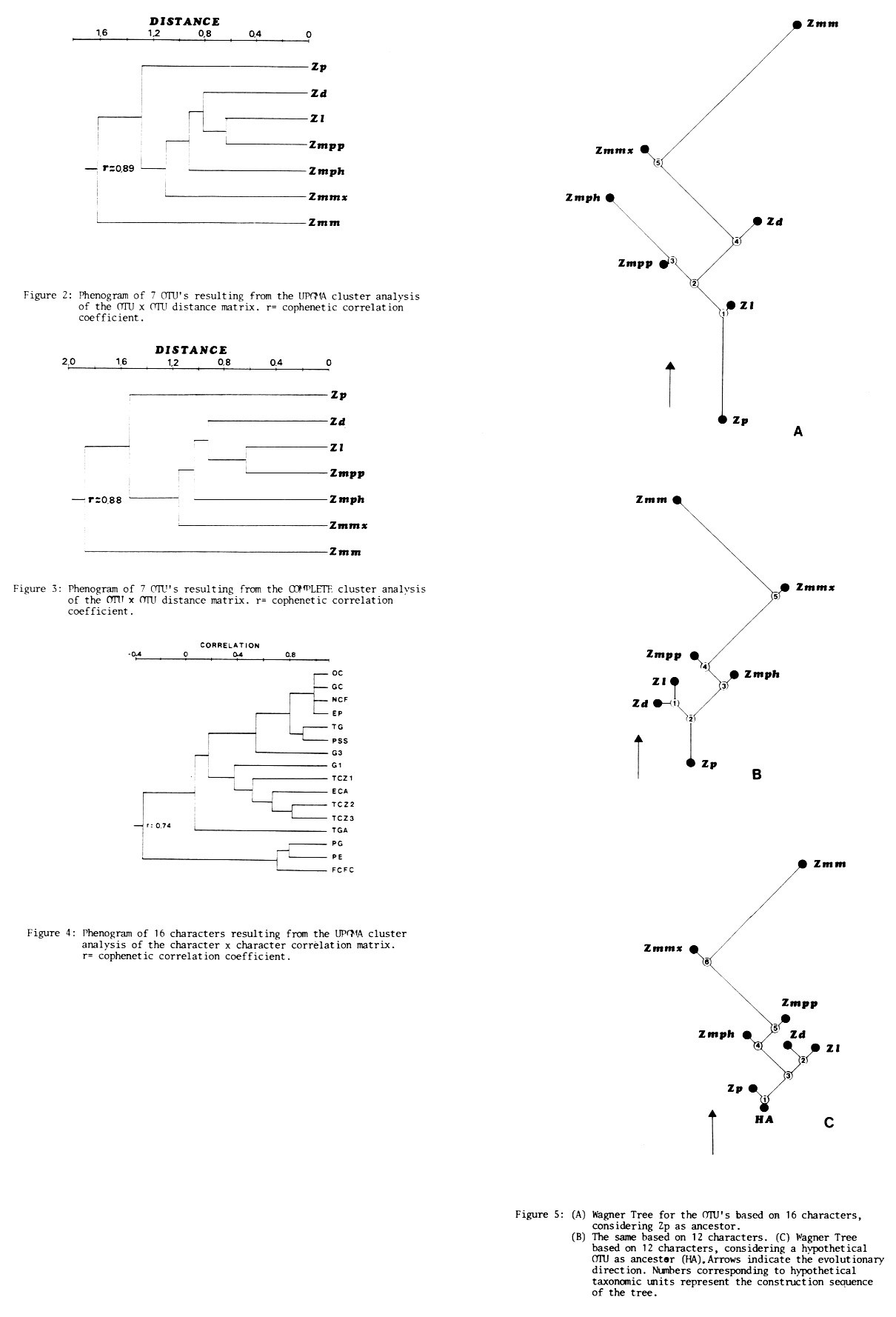
The results obtained from Zea taxa cluster analysis based only on fruit traits appear to be highly congruent with clustering based on other characters (from plant, inflorescence, etc.) reported (MNL 59:61, 1985). Congruence measured by correlation coefficient was high and extremely significant (r = 0. 83). This fact points out the high fruit trait taxonomic value, that allows the taxa to be grouped in an approximately similar way to those done on the basis of more complex studies. Phenograms of Figures 1, 2 and 3 based on fruit traits show that Zp and Zmm are the most extremely differentiated taxa and that Zmpp and Zmph are more like Zd and Zl than Zmm.
Many traits are significantly associated and these traits are those which have most specific value. On the other hand, other traits are represented as isolated by the phenogram shown in Figure 2, and are weakly associated with the remainder, being perhaps, as was pointed out previously (MNL 59:61, 1985), of no specific importance and of little adaptive value. The evolutionary trees of Figure 5 are approximately similar between them. However a fundamental topic is found when choosing those traits in which "Wagner trees" construction is based. Using the 16 characters (Figure 5 A) a certain distortion exists if we consider only 1 to 12 traits (Figure 5 B and C). This happens because those traits which have suffered evolutionary changes and have adaptive value are to be previously stated clearly, removing those traits such as 13 to 16 that don't demonstrate specific value (see MNL 59:61, 1985).
Evolutionary trees built on the basis of 12 fruit traits with evolutionary significance keep a high similitude between them and a high congruence with those built on the basis of 28 traits (see MNL 59:61, 1985). Evolutionary trees show Zp as the most primitive teosinte (considering Zp as ancestral as well as a hypothetical ancestor) to which the other two taxa of Luxuriantes Section (Z. diploperennis and Z. luxurians) link at the lower part of the tree. Balsas teosinte and Huehuetenango are nearer to the most primitive teosintes than to Z. mays ssp. mexicana which is, in accordance with fruit traits, the teosinte more similar to maize.
Those demands, that Wagner's tree (monophyletic) has in its construction, make Zmm linked to Zmmx and placed at the top of the tree. This ought not to be interpreted as if maize had been originated from Zmmx, but as it is the most similar taxon because of being the teosinte with more maize introgression.
Based on our previously discussed hypothesis (MNL 59:61, 1985) it can be pointed out that fruit traits considered isolated may also support the idea that maize introgression, first into most primitive teosintes and then into their derived products, has been the principal factor in teosinte evolution, in the same way as it has played an important role in modern maize evolution.
Table 1. Basic data mtrix (BDM) : Fruit traits.
J.L. Magoja, L.M. Bertoia, I.G. Palacios and M.E.
Streitenberger
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}