In this report the main previous contributions that support the hypothesis concerning the tetraploid nature of Zea mays are reviewed. On the other hand, new cytological evidences supporting a basic number x = 5 for the polyploid complex genus Zea are discussed.
Darlington (Chromosome Botany, Allen & Unwin, London, 1956) considered the concept of basic number and pointed out that the inference of a basic number in a polyploid series is obviously an important step in fitting chromosome numbers to an evolutionary hypothesis. Sometimes the diploid members of the series have disappeared from the genus. At the same time the internal evidence of polyploidy may have disappeared also. According to Darlington "this is the situation in Zea. The ten chromosomes of the haploid set in Zea mays show no internal relations, no evidence of an earlier doubling. Zea mays is what we call and rightly call for all practical purposes, a diploid species". Nevertheless relatives, both in Coix and Sorghum, have the haploid number of 5. This was a strong suggestion, therefore, that 5 is the ancestral basic number (Darlington, ibid.).
Cytological evidences of internal relations among the 10 chromosomes of the haploid set of maize were found by several authors. McClintock (Zeitschr. Zellf. u. mikr. Anat. 19:191-237, 1933) studied prophase pairing in a maize monoploid and reported the occurrence of a high degree of pachytene pairing which looked as intimate as homologous pairing. Chaganti (The Bussey Inst., Harvard Univ., 1965) made a detailed study of various types of chromosome associations in meta-anaphase I in haploid maize and 30% of the cells showed associations implying homology or homoeology. Similar associations have been found recently by Ting (Maydica 30:161-169, 1985) in the first meiotic prophase of haploid maize obtained from anther culture. Vijendra Das (Cytologia 35:259-261, 1970) found secondary association of bivalents in Zea mays (2n = 20) and a maximum of five groups of two trivalents were observed at diakinesis. Finally, the 3-D reconstructions study, using electron-micrographs of serial thin sections of somatic metaphase cells, made by Bennett (Kew Chromosome Conference II, Allen & Unwin, London, pp. 71-79,1983), showed that the 20 chromosomes of Zea mays form four subsets of 5 chromosomes rather than two sets of 10. As Bennett (ibid.) pointed out, his model may have predicted intraspecific homoeology, suggesting that Zea mays is a tetraploid and indicating which chromosomes are homologues.
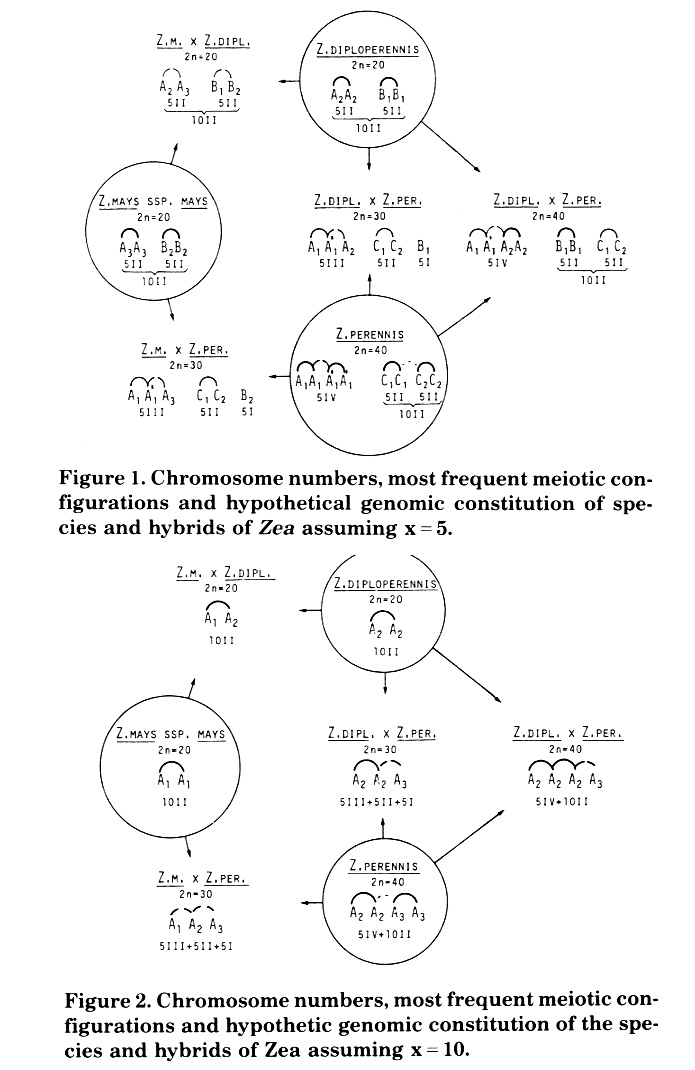
New cytological evidence supporting x = 5 has been obtained as a consequence of our analysis of the meiotic configuration of Zea mays subsp. mays, Z. diploperennis, Z. perennis and of four F1 artificial interspecific hybrids. In Figure 1 the most frequent meiotic configuration in each taxon is presented. The majority of the results could be interpreted only assuming x = 5. Hypotheses about the genome constitution of each taxon are proposed.
In Zea mays subsp. mays (2n = 20, Figure 1) 10 bivalents are regularly formed. Secondary associations were observed and a maximum of five groups of two bivalents was frequent at diakinesis-metaphase I. The meiotic configurations are those of a typical allotetraploid.
As was pointed out by Molina (The Nucleus 26:1-7, 1983; MNL 55:51-52, 1981) Zea diploperennis (2n = 20; Figure 1) presents a regular meiosis with the formation of 10 bivalents in 73% of the cells studied. The formation of 1 III + 8 II + 1 I in 4% of the cells would indicate the segmental allotetraploid nature of this taxon. Secondary associations were also found in this species and groups of four or five pairs of bivalents are frequent.
Zea perennis (2n = 40, Figure 1) had 10 II + 5 IV in 55% of the cells studied (Molina, MNL 56:96,1982; Genetica lberica, in press). These configurations suggest the existence of at least three different genomes (A1, C1, C2). The formation of 5 IV in 55% of the cells would indicate the existence of four highly homologous genomes (A1A1A1A1). The cells with less than 5 IV (4, 3, 2, or 1 IV) probably result from lower chiasmata frequency and do not show formation of III and I. These results suggest high homology by pairs in A1 genomes (A1A1 and A1A1'). Sometimes 6 IV were observed and this could indicate some homologies between C1 and C2 genomes. On the basis of these results Z. perennis would be considered as an auto-allooctoploid with the genomic formula A1A1A1'A1' C1C1C2C2.
The F1 hybrid Zea mays subsp. mays x Z. diploperennis (2n = 20, Figure 1) presents the formation of 10 bivalents in ca. 70% of the 181 cells studied; the rest of the cells present 2 to 6 I (Rosales & Molina, MNL 57:63, 1983). These results suggest that there is exclusive allosyndetic pairing (A2A3 and B1B2). Secondary associations were observed and a maximum of three groups of two bivalents each at diakinesis-metaphase I was formed.
In the F1 Z. mays subsp. mays X Z. perennis (2n = 20, Figure 1) 5 III + 5 II + 5 I formed in ca. 55% of the 80 cells studied (Molina, Rev. Agr. La Plata 56:513-519, 1978). The 5 III could be formed by autosyndetic pairing of A1 and A1' genomes from Z. perennis and by homoeologous pairing with the A3 genome from Z. mays. The 5 II could be formed by autosyndetic pairing between the C1 and C2 homoelogous genomes from Z. perennis. Finally the 5 I would belong to the B2 genome from Z. mays. The same considerations could be applied in the F1 Z. diploperennis x Z. perennis (2n = 30, Figure 1) interpretations. Configurations of 5 III + 5 II + 5 I were present in ca. 40% of the 168 cells studied. In this hybrid the 5 1 could be formed by the B1 genome from Z. diploperennis.
The F1 hybrid Z. diploperennis x Z. perennis (2n = 40, Figure 1) probably originated by the fertilization of an unreduced egg cell from Z. diploperennis by a normal male gamete from Z. perennis (Molina, Cytologia 50:57-62, 1985). The formation of II + IV was observed in 94% of the 214 cells studied, and III + I were not detected. The most frequent configuration, 5 IV + 10 II, was observed in ca. 32% of the cells. These results suggest high homology between A1 and A1 genomes, which is also observed in Z. perennis.
The interpretation of the meiotic configurations of the three species and the four hybrids is more difficult as a whole, if a basic number x = 10 is considered (Figure 2). The present analysis constitutes strong evidence in favor of x = 5 for the genus Zea.
Figure 1. Chromosome numbers, most frequent meiotic configurations and hypothetical genomic constitution of species and hybrids of Zea assuming x = 5.
Figure 2. Chromosome numbers, most frequent meiotic configurations and hypothetic genomic constitution of the species and hybrids of Zea assuming x =10.
Maria del Carmen Molina and Carlos A. Naranjo
Return to the MNL 60 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}