Phylogenetic relationships between maize and its close wild relatives (the teosintes) are a problem closely linked to the origin of maize, which has a century of controversy in its study. In spite of the quantity of hypotheses that have been suggested during this time, basically two schools defending opposite ideas in regard to maize origin were formed: 1) George Beadle's, that maize derives from teosinte, and 2) Paul Mangelsdorf's, that a wild maize is the ancestor of the cultivated one. The aim of this article is to present new proofs or new ways of setting in order the existing data.
Morphological traits: As laid down in the last taxonomic treatment (Doebley and Iltis, Amer. J. Bot. 67:1980), Zea has the following taxa: Z. perennis (perennial teosinte) (Zp), Z. diploperennis (diploperennial teosinte) (Zd), Z. luxurians (Guatemala teosinte) (ZI), Z. mays ssp. parviglumis var. parviglumis (Balsas teosinte) (Zmpp), Z. mays ssp. parviglumis var. huehuetenangensis (Huehuetenango teosinte) (Zmph), Z. mays ssp. mexicana (NobogameChalco-Central Plateau teosinte) (Zmmx), and Z. mays ssp. mays (maize) (Zmm). These constitute the 7 operational taxonomic units (OTU's).
Ten morphological traits were scored for each OTU as follows: (1) tassel central spike distichous (0), polystichous (2); (2) Cupulate fruit case trapezoidal (2), triangular (1), horizontally compressed (0); (3) Kernel size small (0), intermediate (1), large (2); (4) Number of fruit cases per spike; (5) Female spike distichous (0), polystichous (1); (6) Female spikelet arrangement single (1), paired (2); (7) Growth habit perennial (1), annual (0); (8) Rhizomes present (1), absent (0); (9) Chromosome number 2n = 40 (2), 2n = 20 (1); (10) Chromosome knob position terminal (1), terminal and internal (2). Given a basic data matrix (BDM) (Table 1) of 10 characters by 7 OTU's, the data were analyzed by cluster analysis. The BDM was standardized (BDMS) by characters to remove the unequal weights imposed on it by the use of different scales of measurement.
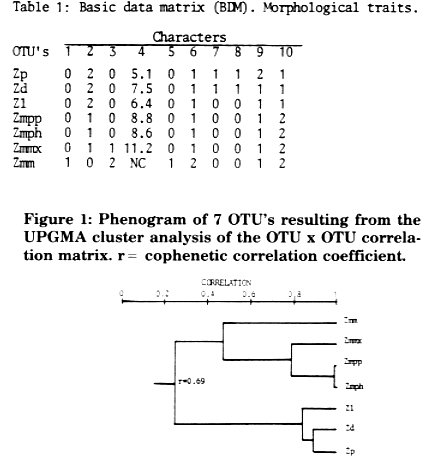
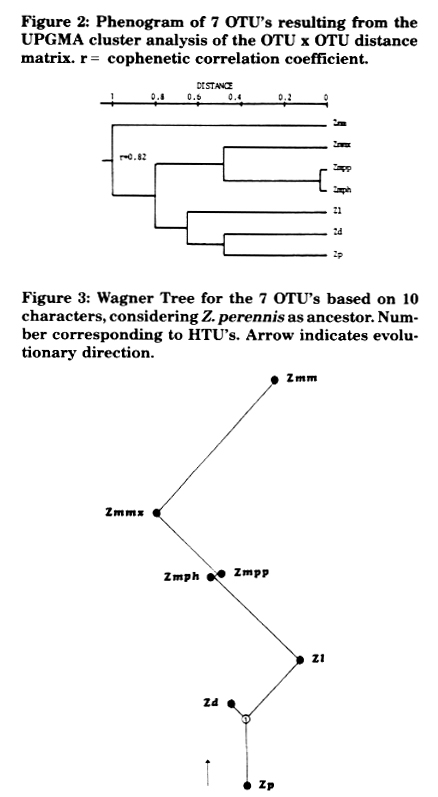
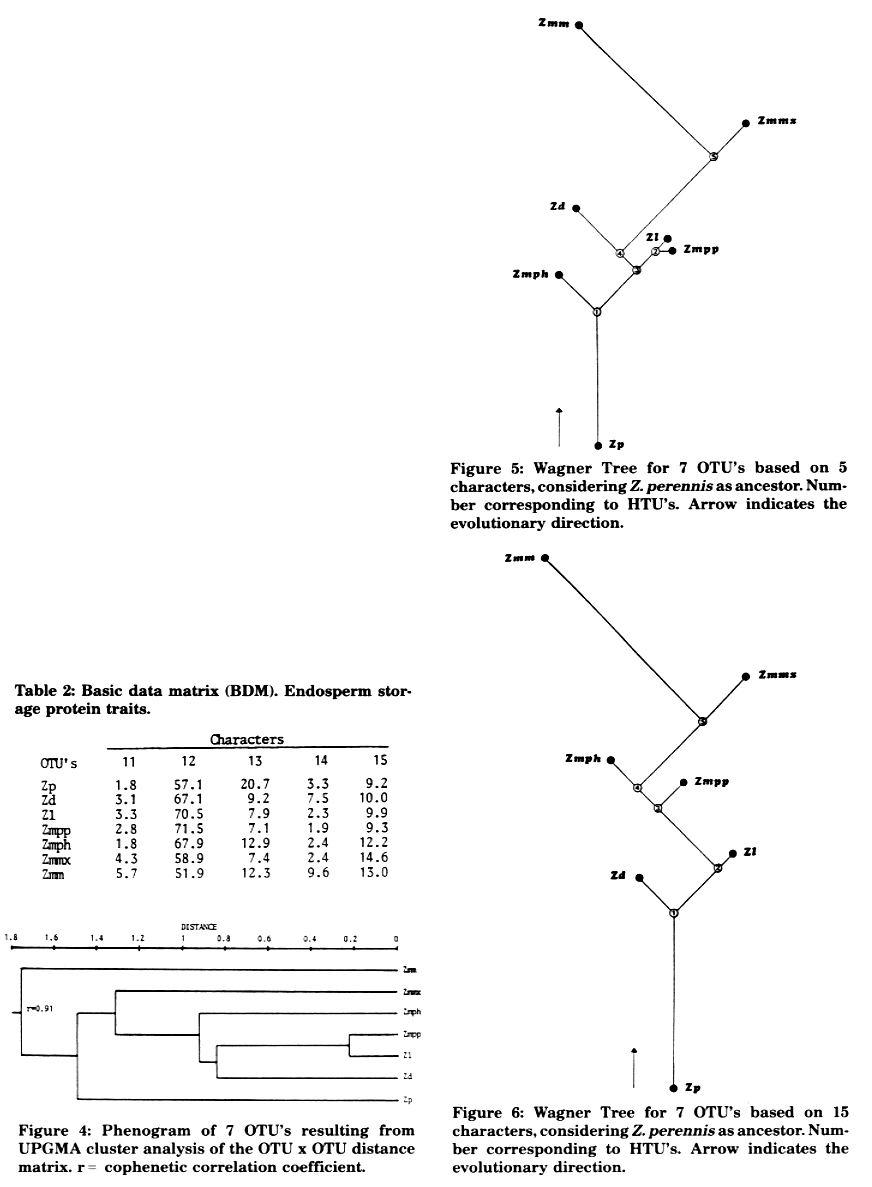
Phenograms were derived by (1) the Pearson product-moment correlation coefficient, applying the unweighted pair group method and arithmetic averages (UPGMA) (Figure 1), (2) the "Mean Taxonomic Distance" between pairs of OTU's (Figure 2), and (3) the "Manhattan Distance" between pairs of OTU's to calculate a "Wagner Tree" (Figure 3). Methods (1) and (2) gave similar phenograms, coinciding with Doebley and Iltis' new taxonomy separating section Luxuriantes (Zp, Zd, Zl) from Zea (Zmm, Zmmx, Zmpp and Zmph); the cophenetic correlation coefficient of -0.82 indicates a lower distortion in method (2), where Zmm segregates further from the others. The tree derived by method (3) was constructed by selecting Zp as ancestor because, in our opinion, it has the largest quantity of morphological characters in a primitive state: it is perennial, has rhizomes, trapezoidal fruit cases and the lowest number of fruit cases per spike. It does not present, unlike Z. diploperennis, an exuberant development, perhaps because it is the only teosinte which has not been contaminated with maize germplasm. Zd and Zl are derived based on a hypothetical taxonomic unit (HTU1) which, together with Zp, constitute the most primitive teosinte group placed at the bottom of the tree. Zmph, Zmpp and Zmmx, of which the first two are very close together, are clustered in the medium level of the tree. At the top of the tree, and very far from the other OTU's, is placed Zmm. Wagner's tree index of consistency is high (C = 0.91) and points out few extra evolutionary processes. The cluster of the taxa presented, based on few morphological characters and using different methods (cluster analysis and Wagner's tree), comes out as highly concordant.
In our opinion the key to mechanisms that have conditioned Zea evolution lies in maize (Zmm) and perennial teosinte, two basic species (and the most distant) that gather all necessary genetic information to explain the other taxa differentiation. The greatest part of individuals in hybrid progeny between perennial teosinte and maize are highly heterotic. In accordance with their morphology, they present phenotypes that move away not only from parental ones but also from those that might be expected as a consequence of their combination. These types of individuals may have distichous spikes with single spikelets, tenacious rachis and enclosed kernels. They may also have distichous spikes with badly enclosed kernels and fragile rachis, as well as any other possible combinations of these characteristics. There is no doubt that hybrids between Zp and Zmm individuals are almost completely inadequate to perpetuate in a natural environment. This failure to adjust prevents them from perpetuation without human aid. This is a consequence of the loss of wild characters, which has taken place with maize germplasm introgression. The maladjustment originates in the incapacity to spread their kernels freely, and because their female inflorescence structure (half-naked kernels not protected by husks) makes them vulnerable to insect attack, diseases, and birds and other animal injuries. Two principal facts demonstrate that maize introgression into primitive teosintes led to evolved teosinte differentiation: a) triangular-fruit-case individuals appear in hybrid progeny, resembling annual teosintes; b) maize germplasm introgression into teosinte with a low number of fruit cases per spike conditions an increase in their number (more-evolved annual teosintes also have this characteristic). Based on tassel characters (neutral characters), it can be stated that maize introgression into more primitive teosintes conditions the phenotype formation with characteristics of the most evolved group.
All things considered, it can be stated that there are enough morphological proofs to hold up the hypothesis that the different teosintes are products of maize germplasm introgression (followed by natural selection), first in perennial teosinte and secondly in the products derived from this first introgression. Consequently, if Z. mays ssp. mexicana is a lot like maize as a result of the maize germplasm accumulation, the cultured species could have never been derived from it. Maize should derive from other species (perhaps a primitive maize in accordance with Mangelsdorf) and, as can be demonstrated experimentally, maize has performed a fundamental role in teosinte evolution.
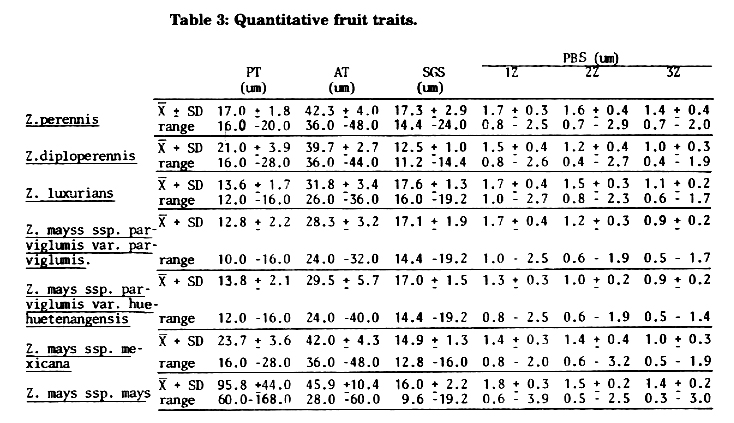
Endosperm storage protein traits: In previous reports (MNL 55:62,1981; MNL 56:106,1982; Magoja et al. Rev. Fac. Agron. 3:255, 1982) we stated that endosperm protein pattern obtained according to Landry-Moureaux's technique could be considered as a specific characteristic that may be used to distinguish maize from its wild relatives. We inferred that maize and perennial teosinte protein patterns were basic and highly different, and intermediate for Z. diploperennis and Z. mays ssp. mexicana (according to the results of other authors), although up to 1982 protein patterns only were studied for Z. perennis, Z. diploperennis and several maizes. The hypothesis that intermediate protein patterns appearing in the other taxa of Zea might derive from that of Z. perennis (primitive protein pattern) by maize germplasm introgression was formulated. Now we present evidence that ratifies this hypothesis. A sufficient amount of annual teosinte seed was generously sent by Hugh H. Iltis in the beginning of 1984, allowing us to start biochemical analysis and to complete protein pattern study in each taxon of Zea. Five Landry-Moureaux endosperm storage protein traits were scored for each OTU as follows: (11) saline soluble proteins; (12) zein; (13) glutelin-1; (14) glutelin-2; (15) glutelin-3. Given a basic data matrix (Table 2), "Mean Taxonomic Distance" served as input in the calculation of a phenogram by the UPGMA (Figure 4), and "Manhattan Distance" in the calculation of a "Wagner Tree" (Figure 5).
Reasonable congruity exists between evolutionary trees based on morphological characteristics and on protein patterns: Zp is situated at the bottom of the tree, Zmm on the top, Zmmx looks like maize. The other taxa are located in medium positions between Zp and Zmmx.
Finally, an evolutionary tree, such as represented in Fig. 6, may be built considering morphological and protein characteristics jointly. This tree has at the bottom Zd and ZI in addition to Zp (the ancestral species), forming the most primitive group of taxa (sect. Luxuriantes, Doebley and Iltis, 1980). In the medium level appear Zmph, Zmpp and Zmmx (sect. Zea), the first two taxa nearer to Zl than to Zmmx. Maize (Zmm) is placed far away from the rest of the taxa, at the top of the tree. The index of consistency of the tree is high (C = 0.77).
Based on the hypothesis that maize introgression into Z. perennis was the predominant factor in Zea evolution, it can be stated, in accordance with morphological and protein traits, that the evolutionary tree shown in Fig. 6 represents what might have occurred in nature: a former maize introgression into Zp led first to Zd (a species that retains perennialism) and then to Zl (annual) differentiation. Consecutive introgressions into Zl led to Zmpp and Zmph formation, which are (except Zl) the annual teosintes closest to the primitive group (ZI-Zd-Zp). Zmmx was produced by the latter maize introgression, and is the most evolved teosinte, because it resembles maize, both in morphological and in protein characters. In short, Zmmx is a maize that retains from teosinte only the essential genes for wild life, turning out to be, consequently, the teosinte with the largest quantity of maize germplasm.
Fruit traits: Four quantitative characteristics of grain were evaluated: pericarp thickness (PT), aleurone layer thickness (AT), starch granule size (SGS) and zein or protein body size (PBS) in three endosperm zones (See MNL 58:129). PT and AT were measured in the abgerminal side of the kernel, SGS measurement was made in the 10th cell layer counting from the aleurone layer downwards inside the grain. All the measurements were made upon three cuts of each kernel, using ten grains taken at random from each species.
The results corresponding to maize are the average of measurements obtained over several types whose PT and AT are different (Table 3). PT is lower in teosinte than in maize. Zmmx has the thickest pericarp of all teosintes. AT is lower in teosinte than in maize, although in maize the aleurone layer thickness is quite variable. SGS seems to have no specific relationship, and no significant differences were found among the diverse taxa. The same can be pointed out for protein body size, which is quite similar in the different taxa of Zea.
The quantitative characteristics were assessed for correlations, as well as other characteristics of these fruits, such as kernel presentation (KP), cupulate fruit case shape (CFCS), grain size (GS), endosperm protein content (EP), saline soluble protein content (SS) and zein content (Z) (Table 4). Some fruit characteristics are significantly associated: grains enclosed in fruit cases are small or intermediate, have a low rate of SS, and their pericarps are thin. Trapezoidal-fruit-case teosintes have the smallest grains. They have higher endosperm protein content, low SS content, high Z content and thinner pericarp. Those taxa with higher endosperm protein content have more zein and thinner pericarp. Those taxa with higher SS content have thicker pericarp. Zein content is higher in those taxa with a thinner aleurone layer and zein bodies are smaller in zone 3. Zein body sizes in zones 2 and 3 are positively associated. From all the characteristics evaluated only pericarp thickness and aleurone layer may be considered specific.
In general, specific characteristics are significantly associated between themselves and with other specific characteristics considered before, such as cupulate fruit case shape, kernel size, saline soluble protein content and zein content. Only endosperm protein content, pericarp thickness and aleurone layer may be considered specific among all the new characters presented in this work. The other ones are very variable in each taxon, and no significant differences are manifested.
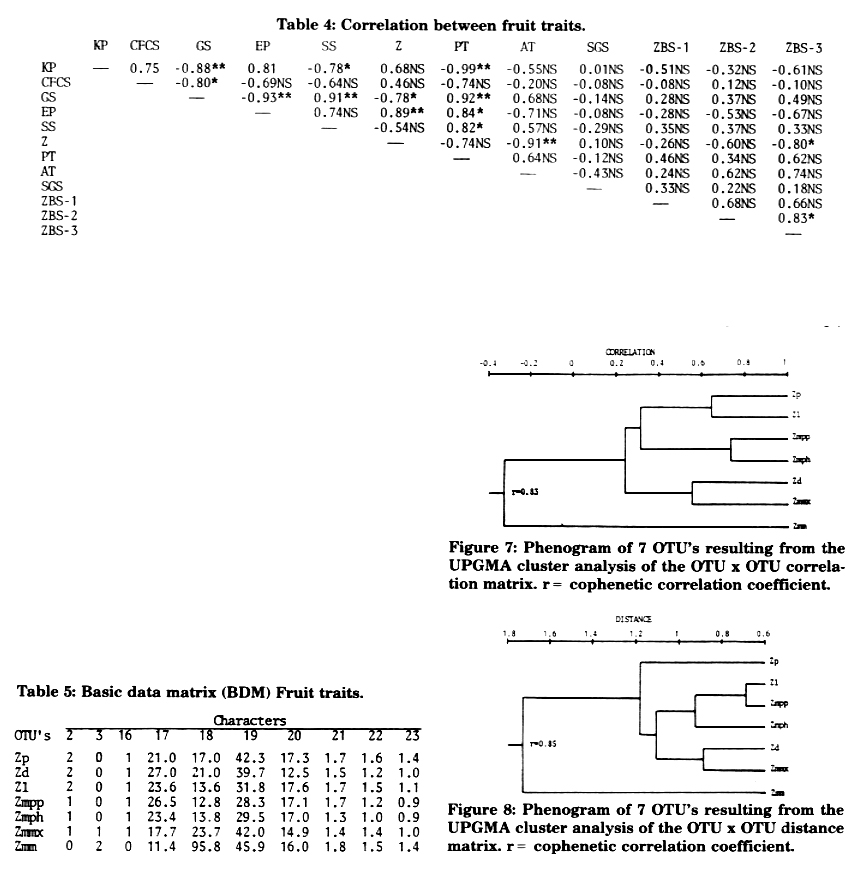
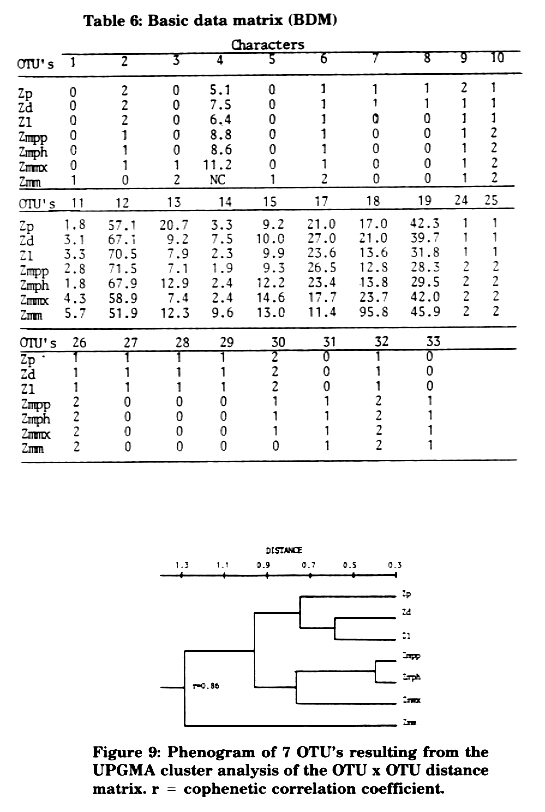
Cluster analyses were performed on fruit data from 10 characters (3 qualitative and 7 quantitative) of taxa of Zea, as follows: (2) Cupulate fruit case shape trapezoidal (2), triangular (1), horizontally compressed (0); (3) Kernel size small (0), intermediate (1), large (2); (16) Kernels enclosed (1), naked (0); (17) Kernel protein content (%); (18) Pericarp thickness (Rm); (19) Aleurone layer thickness (µm); (20) Starch granule size (µm); (21) Zein body size of zone 1 (µm); (22) Zein body size of zone 2 (µm); (23) Zein body size of zone 3 (µm). Given a basic data matrix (BDM) (Table 5), the Pearson product-moment correlation coefficient served as input in the calculation of a phenogram by UPGMA (Figure 7), and "Mean Taxonomic Distance" between pairs of OTU's was calculated (Figure 8). In both phenograms Zmm links at the farthest distance, which means it is the most differentiated species in regard to fruit characteristics. There is no high congruency in clustering based only on fruit characteristics, as observed in those based on other characteristics. This may occur because of having included some non-specific fruit characteristics (which have been pointed out before) with the same weight as the specific ones according to the exigencies of the numerical method chosen.
In accordance with the results obtained, it can be determined that fruit characteristics can not be used separately to assemble the taxa of Zea, and that from the characteristics studied, protein content, pericarp thickness and aleurone layer are the most specific ones with evolutionary significance.
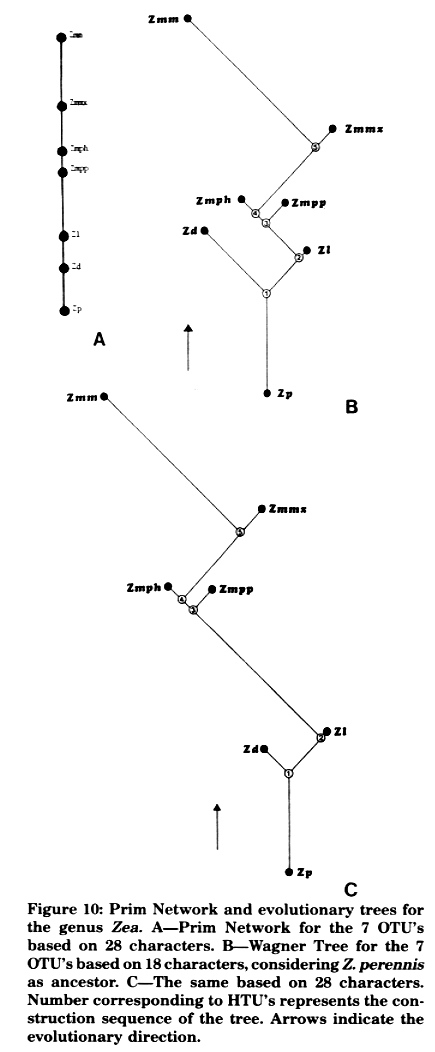
Analysis of dispensable traits: In addition to the characteristics employed in previous articles, we also used "neutral characters" or tassel traits according to Doebley and Iltis (1980). Ten neutral morphological traits were scored for each OTU as follows: (24) Tassel branch number low (1), high (2); (25) Tassel branching axis short (1), long (2); (26) Lateral tassel branch internode short (1), long (2); (27) Male spikelet outer glume wing absent (0), present (1); (28) Male spikelet outer glume primary lateral vein narrow and scarcely (0), wide and prominent (1); (29) Male spikelet outer glume total veins few (0), numerous (1); (30) Tassel branching abscission layer absent (0), strongly to weakly developed (1), strongly developed (2); (31) Tassel branching rachis flattened (0), rounded (1); (32) Male spikelet outer glume stiff (1), papery (2); (33) Male spikelet outer glume flattened (0), rounded (1). Given a basic data matrix (BDM) (Table 6) of 28 characters by 7 OTU's, the data were analyzed by cluster analysis. The BDM was standardized (BDMS) and data were analyzed by "Mean Taxonomic Distance" (Figure 9), by "Manhattan Distance" (Figure 10) and a "Prim Network" (Figure 10A), and by "Manhattan Distance" and "Wagner Trees" (Figure 10B, C).
Figure 9 shows a phenogram with 2 definite clusters: one of them constituted by the most primitive teosintes (Zp-Zd-ZI) and the other by the most evolved ones (Zmpp-Zmph-Zmmx). Maize (Zmm) segregates separately from the rest of the taxa, and is located as distant from one group as from the other. The Prim Network given in Figure 10A shows that Zp and Zmm are the two taxa situated at the farthest distance from each other, whereas the most primitive teosintes link nearer to Zp than the most evolved ones. The latter are located between ZI and Zmm. Wagner Trees shown in Figure 10B and C are similar. High congruity exists whether 18 traits (1 to 10 orthodox morphological traits + 11 to 15 endosperm protein traits + 17-18-19 fruit traits; see previous reports), or 28 traits including "neutral morphological characters", are considered. The taxa ordering is similar in both trees and coincidental with the Prim Network. Results obtained point out that evolutionary trees constructed with either 18 or 28 characters have a high index of consistency (C = 0.78 and C = 0.82, respectively), denoting a few extra evolutionary processes. Both phenograms (Fig. 9) and evolutionary trees (Fig. 10) clearly demonstrate an extremely particular and well defined clustering: teosinte grouped in 2 levels (one primitive and the other evolved), and maize separated from them.
Results obtained in cluster analysis strongly uphold that taxa of Zea are the product of maize introgression, first in the most primitive teosinte (Zp) and secondly into the derived taxa. Introgression degree was increased at the same time as the tree is climbed, until it reaches Zmmx, the teosinte with the maximum maize introgression. If Zmmx is the teosinte with the highest maize introgression, as can be supported by morphological, biochemical and cytological evidence, it could never have been the maize ancestor.
When this series of reports about "Evolution of Zea" was initiated in this MNL, it was stated that our purpose was that new data or new ways of ordering the existing data would be provided, in order to clarify phylogenetic relationships among the taxa of Zea, reinforcing an old hypothesis. Now, we want to explain that these new data and the new way of ordering them come from our own research, and this old hypothesis to which we refer is Paul Mangelsdorf's.
Table 1: Basic data matrix (BEM). Morphological traits
Figure 1: Phenogram of 7 OTU's resulting from the UPGMA cluster analysis of the OTU x OTU correlation matrix. r = cophenetic correlation coefficient.
Figure 2: Phenogram of 7 OTU's resulting from the UPGMA cluster analysis of the OTU x OTU distance matrix. r = cophenetic correlation coefficient.
Figure 3: Wagner Tree for the 7 OTU's based on 10 characters, considering Z. perennis as ancestor. Number corresponding to HTU's. Arrow indicates evolutionary direction.
Table 2: Basic data matrix (BDM). Endosperm storage protein traits.
Figure 4: Phenogram of 7 OTU's resulting from UPGMA cluster analysis of the OTU x OTU distance matrix. r = cophenetic correlation coefficient.
Figure 5: Wagner Tree for 7 OTU's based on 5 characters, considering Z. perennis as ancestor. Number corresponding to HTU's. Arrow indicates the evolutionary direction.
Figure 6: Wagner Tree for 7 OTU's based on 15 characters, considering Z. perennis as ancestor. Number corresponding to HTU's. Arrow indicates the evolutionary direction.
Table 3: Quantitative fruit traits.
Table 4: Correlation between fruit traits.
Table 5: Basic data matrix (BDM) Fruit traits.
Figure 7: Phenogram of 7 OTU's resulting from the UPGMA cluster analysis of the OTU x OTU correlation matrix. r = cophenetic correlation coefficient.
Figure 8: Phenogram of 7 OTU's resulting from the UPGMA cluster analysis of the OTU x OTU distance matrix. r = cophenetic co"elation coefficient.
Table 6: Basic data matrix (BDM)
Figure 9: Phenogram of 7 OTU's resulting from the UPGMA cluster analysis of the OTU x OTU distance matrix. r = cophenetic correlation coefficient.
Figure 10: Prim Network and evolutionary trees for the genus Zea. A-Prim Network for the 7 OTU's based on 28 characters. B-Wagner Tree for the 7 OTU's based on 18 characters, considering Z. perennis as ancestor. C-The same based on 28 characters. Number corresponding to HTU's represents the construction sequence of the tree. Arrows indicate the evolutionary direction.
J. L. Magoja, I. G. Palacios, L. M. Bertoia and M. E. Streitenberger
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}