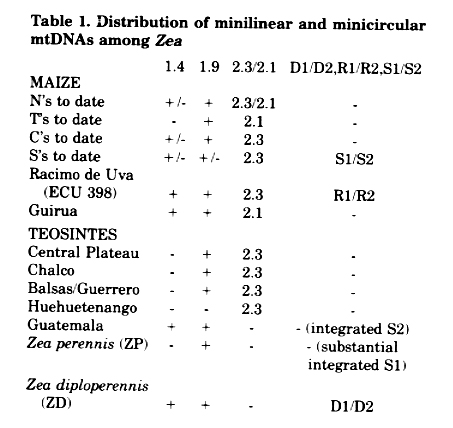
Maize and its relatives share a number of small minilinear or minicircular mtDNAs, ranging from ca. 7.4 to 1.4 kb. An inspection of recent literature suggests at least 20 such small molecules; common among these are the D1/D2, R1/R2, S1/S2, and 2.3/2.1 kb minilinear DNAs and the 1.9 and 1.4 kb minicircular DNAs. As part of investigations of mtDNAs in maize and teosinte, we have assembled a limited compilation of occurrence of these extraprincipal-genome molecules. Occurrence of the DNAs was determined by probing with cloned S1/S2, 1.9, and 1.4 kb molecules, and by visual inspection of DNAs for the 2.3/2.1 kb minilinear DNAs. The data suggest an element of randomness of distribution within Zea.
Our survey (Table 1) is limited in extent, but nevertheless certain trends are indicated. We have examined at least 15 normal cytoplasms, two entries of T cytoplasm, five members of the C group, and nine members of the S group. The teosintes utilized were the same as previously described (Timothy et al., Proc. Natl. Acad. Sci. 76:4220, 1979). One seed lot of Zea diploperennis (designated 540), one seed lot of RU (ECU 398), and the Latin American race Guirua, were provided by D. H. Timothy, NC State, Raleigh.
The 1.4 kb minicircular DNA is limited in distribution, present in some N's, C's, and S's, and in only Guatemala and Zea diploperennis. The 1.9 kb minicircular DNA is more widely distributed, but absent in some S's and in Huehuetenango. However, sequences of this molecule appear to vary in size in the teosintes, RU, and Guirua. This variation is reflected in apparent kb differences of undigested and restriction endonuclease-digested samples, with most bearing slightly larger molecules than the 1.9 kb molecule.
The 2.3/2.1 kb minilinear DNA is absent in only Guatemala, ZD, and ZP in entries examined to date. The 2.1 replaces the 2.3 kb DNA in T's, some N's, and in the Latin American race Guirua. An accession designated Puya Grande also carries the 2.1.
A number of these mtDNAs were probed with the three PstI clones which comprise most of the S2 DNA, an internal 1.55 kb HindIII fragment of S1, and a 6.2 kb PstI clone of S1, to survey the frequency of homologies to S1/S2 sequences in the principal genome. All normal cytoplasms examined to date and the teosintes Central Plateau, Chalco, Balsas/ Guerrero, Huehuetenango, and ZP are similar and characterized by an apparently small deletion at the end of the S2 sequence which shares ca. 1500 bp homology with S1. Guirua has only trace homology to S2, which is unique among apparently normal, male fertile cytoplasms, and is a characteristic of the T and C cytoplasms. Guatemala apparently carries a total integrated copy of S2, as evidenced by intactness of the three PstI fragments which comprise most of the molecule; the only other maize which is similar to date is S cytoplasm. Even though ZP is like normal cytoplasm maize in the S2 region, ZP may carry a large integrated part of the S1 molecule. One major BamHI fragment and at least two large PstI fragments carry homology to the S1-specific HindIII 1.55 kb clone and to the PstI 1.6 kb clone which includes most of the ca. 1500 sequence shared by S1/S2. A complete copy of S1 was not detected by virtue of absence of a 6.2 kb PstI fragment which hybridizes to these two clones. Nevertheless, no other Zea cytoplasm examined to date except S yields evidence of integration of S1 to this extent.
Guatemala, ZP, and ZD, at least our seed lots, display unique and diagnostic endonuclease restriction patterns, but do share some pattern affinity. These patterns are distinct from other teosintes (Timothy et al., Proc. Natl. Acad. Sci. 76:4220, 1979) or other normal cytoplasms (Sederoff et al., Proc. Natl. Acad. Sci. 78:5953, 1981). ZD carries the free D1/D2 DNAs, which are similar to the R1/R2 DNAs, which may be progenitors of S1/S2. Although the significance of these observations is unclear at present, these teosintes may be intimately associated with the evolution of S1/S2 sequences. The three teosintes are unique in that they are the only Zea entries examined to date that lack the 2.3/2.1 minilinear DNAs. It is also interesting to note that defective endosperm is associated with the ZP cytoplasm in certain lines, and that Guatemala cytoplasm is also associated with a small seed trait.
These observations suggest randomness of distribution and independent evolution of extra-principal-genome mtDNA sequences in Zea. Sequences of these molecules appear to be dispensable to the genus, in that one entry or another may lack all of these sequences. However, an intimate role of S2 sequences, or more likely S2/R2/D2 sequences, in mtDNA rearrangements associated with reversion to fertility in the S cytoplasm is suggestive of involvement in the evolution of Zea mtDNA. That Guirua, T, and C cytoplasms share very little homology with S1/S2 indicates that these sequences, too, are not universally present in Zea.
Table 1. Distribution of minilinear and minicircular mtDNAs among Zea.
D. R. Pring and A. G. Smith
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}