This report is concerned mainly with a new look at data previously reported in MNL or elsewhere. In two publications (1958, 1974), I suggested that a mutation system that had originated in my hybrids of corn and teosinte had features in common with those of McClintock (1950), Brink (1951), and others. An article by Nina Fedoroff in the June number of Scientific American has revived my long-dormant interest in mutation systems and prompted me to reexamine the results of my earlier experiments, especially to see if they could explain the source of the mobile genetic elements now known as transposable elements.
In the cytogenetic research on corn and its relatives in which Robert Reeves and I were engaged from 1927-1940, we found that annual teosinte, corn's closest relative, differs from corn, not by a small number of genes controlling specific morphological traits but by blocks of genes affecting many characteristics. Later in crosses with a nine-gene linkage tester, I found four of these blocks to be located on chromosomes 1, 3, 4, and 9. The tester had no marker-gene for chromosome 5 (MNL, 1947, p.20, 1974, p.41).
To study these blocks in a uniform genetic background, I crossed them with a well adapted Texas inbred 411-3 and by repeated backcrossing produced modified strains differing primarily in the blocks of genes introduced from annual teosinte. When I moved to Cambridge in 1940, I found these modified Texas strains to be poorly adapted to Massachusetts and by another cycle of backcrossing, introduced them into a second inbred, Minn. 158, an early-maturing strain. After four backcrosses to A158, the strains were selfed.
In several of the S1 progenies, mutations occurred at rates higher than those produced by massive doses of x-rays. The mutations were of two kinds, stable and unstable. The latter are of special interest here, one in particular, a defective-seed mutant that inhibits the development of the endosperm in varying degrees from almost none to almost complete, the latter resulting in caryopses with little more than collapsed pericarps.
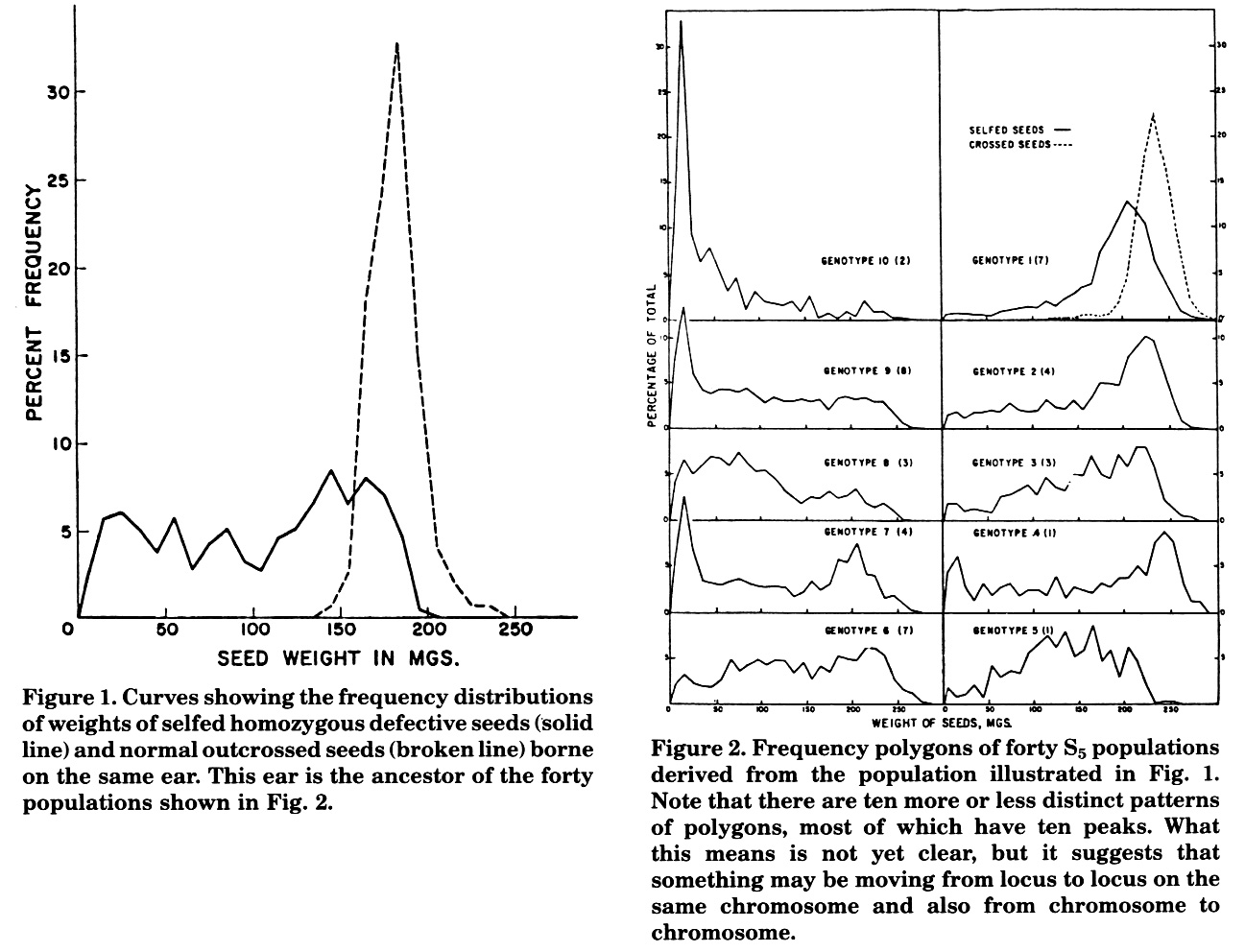
On a doubly-pollinated ear, homozygous for the mutant defective, the weights of the outcrossed seeds ranged from 140 to 250 mg and the frequency curve depicting these was the well-known normal frequency curve with a single conspicuous peak (Figure 1). In sharp contrast was the frequency distribution depicting the weights of the defective seeds. These varied from 25 to 200 mg and their polygon had 5 distinct peaks.
In the next several generations, the peaks depicting the weights of the defective caryopses increased from 5 to 6, 7, 8, 9 and 10. Also, there were "repeats" in the patterns of the polygons. Some pairs were so similar that their peaks coincided almost perfectly.
Finally, in a population of forty S5 ears, there were ten more or less distinct patterns each, with one or two exceptions having ten peaks but no more (Figure 2). Ten happens also to be corn's chromosome number; whether this is more than coincidence is a moot question. What is clear, however, is that the S2 population with 5 distinct peaks has been the ancestral source of a family of populations with numerous patterns of frequency polygons each with numerous peaks. This may suggest that mobile elements of some sort were not only becoming more numerous but may have been moving from chromosome to chromosome, until they became, in Fedoroff's words, "ubiquitous".
In both of my publications (1958, 1974) I called attention to similarities between this mutation system and that of McClintock and others, but offered no convincing explanation of their origin. In the more recent one (1974), I did suggest that chromosomes introduced into corn from teosinte, although appearing by linkage tests and conventional cytological studies to be homologous to those of maize, might not be in perfect register, so that crossing over at various points could produce small deficiencies and duplications. The former might be lethal or inhibiting in their effects; the latter might create useful changes that were preserved under domestication. The possibility that unequal crossing over has been a factor in corn's evolution under domestication should not be ruled out unless studies at the molecular level do so.
Having reexamined my own data, I turned to the more recent results of McClintock and others. Of these the most interesting and perhaps most significant are those set forth in Table 1 in a 1965 paper (McClintock, 1965). This shows that the chromosomes identified as being involved in the Ac, Spm, and Dt mutation systems are 1, 3, 4, 5, and 9. This rings a bell! I am reminded that my 1946 linkage studies of blocks of genes derived from annual teosinte showed these to be located on four of these same chromosomes: 1, 3, 4, and 9. The tester, it may be recalled, had no marker gene on chromosome 5. The odds against even four of the same chromosomes being involved in the two mutation systems by chance alone are considerable. Calculated in one way they are significant; with other valid assumptions, substantially higher, and they are augmented by other circumstances.
All four of the blocks of genes that I had introduced into an adapted inbred strain of corn were associated with "tripsacoid" cobs. These are identified by stiff indurated outer glumes, often curved, that give the cobs the impression and feeling of a wood rasp. In fact it was this characteristic, more than any other, that enabled me to identify the blocks through two cycles of backcrossing.
In Plate I of a 1967 paper (McClintock, 1967) 13 cobs illustrating several types of variegation are depicted. All appear to be tripsacoid. Of almost equal interest is an ear illustrated in Plate 2D of the same paper showing an ear segregating for defective seeds (abortive caryopses).
In view of all of these additional similarities between the two mutation systems, McClintock's and mine, can there still be much doubt that they are in some way related? Fortunately the question can now be answered by studies at the molecular level. I am told that these are now in progress at several laboratories in this country and abroad. I await the results with more than casual interest!
In the meantime it may be useful for me to suggest to those who are engaged in research in this area some of the abundant materials that are available. Especially useful should be the modified inbred strains of A158 that have given rise to mutation systems. Walton Galinat may know what is available in this group. Also useful may be modified strains of A158 that have been introgressed by chromosome 4 of Zea diploperennis, which our experiments have shown to be probably ancestral to the annual teosintes. A small S1 population of A158 in which the gene for perennialism has been introduced by repeated backcrossing segregated in 1983 for defective seeds, dwarfs and male sterility. This is an apparent mutation system tracing directly to the ancestral source rather than to the annual teosintes that represent an intermediate stage.
Fedoroff suggests that some of the variegation in races of maize of Central and South America may represent mutation systems and that it would be of interest to determine if these are similar or different from those already recognized. There is such a system in the Peruvian race Huayleno in which virtually all the pericarp and cob colors and patterns known anywhere in the world occur in one narrow valley in the Department of Ancash (Grobman et al., 1961). Since variegation is not found in the prehistoric ears of corn in Peru, where they are abundant in archaeological sites, it may be assumed that there has been hybridization either with an introduced race from Mexico carrying teosinte germplasm or with a species of Tripsacum of which several occur in Peru. Other Peruvian races of special interest are Cuzco with variegated pericarp and Pisccorunto for its stippled aleurone.
All of the authors of the ten publications on the races of maize of the countries of this hemisphere, beginning with Wellhausen's classic, Races of Maize of Mexico, may enjoy a degree of satisfaction if some of the races that they collected, described, and preserved for utilitarian purposes turn out to be also useful in solving the mystery of the mobile genetic elements popularly known as "jumping genes".
Figure 1. Curves showing the frequency distributions of weights of selfed homozygous defective seeds (solid line) and normal outcrossed seeds (broken line) borne on the same ear. This ear is the ancestor of the forty populations shown in Fig. 2.
Figure 2. Frequency polygons of forty S5 populations derived from the population illustrated in Fig. 1. Note that there are ten more or less distinct patterns of polygons, most of which have ten peaks. What this means is not yet clear, but it suggests that something may be moving from locus to locus on the same chromosome and also from chromosome to chromosome.
Paul C. Mangelsdorf
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}