The loci pale green-11 and pale green-12 in the double recessive homozygote, pg11 pg11 pg12 pg12, determine a pale green plant phenotype. In dominant genotypes, which have a wild phenotype, we have studied, at the quantitative level, differences in the following traits: days from planting to silking and pollen shedding date, number of tassel branches, number and weight of ears, number of nodes below the ear, number of total nodes, ear height and plant height.
From an inbred line with an inbreeding coefficient of 0.996 with origin in ((UR x A545o2)UR2)sel.6, but still segregating at the loci pale green-11 and pale green-12, near isolines Pg11 Pg11 Pg12 Pg12 (1111), Pg11 Pg11 pg12 pg12 (1100), pg11 pg11 Pg12 Pg12 (0011) and pg11 pg11 pg12 pg12 (0000) were separated by means of a wx marker. By selfing and crossing among near isolines, the 8 possible wild genotypes were obtained in a common homozygous genetic background. The same near isolines were crossed with each one of the inbreds MC (0000), W64o2 (0011), URo2 (1100) and B37o2 (1111), and heterozygous F1 progeny were obtained. From these materials and the traits mentioned above comparisons were made among: 1) different homozygous genotypes in a common background, and 2) heterozygous genotypes in different genetic backgrounds.
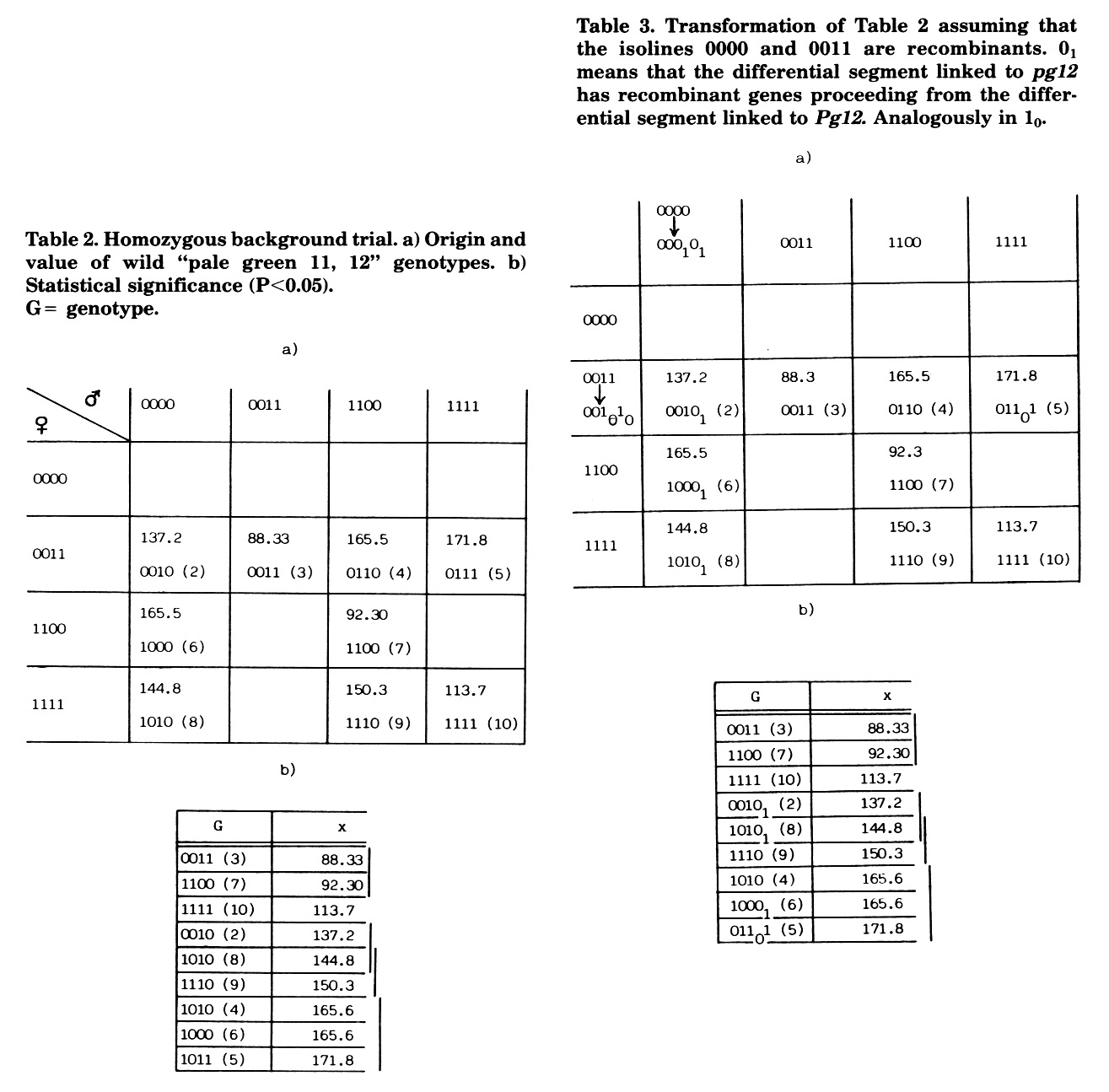
In the homozygous genetic background trial, the genotype Pg11 pg11 Pg12 pg12 (1010) was represented twice, originating from crosses of different near isolines (Table 1). Between these two progenies, significant differences in three traits were found, which can be ascribed to the segregation of general heterozygosity (0.4%) or to genetic recombination in the differential chromosome segments linked to the markers. The recombination event should have occurred in the passage from the original plant to the first generation of near isolines. Assuming that the differential segments linked to the markers arose mostly from UR, if linked to Pg11 or pg12, and from A545o2 if linked to pg11 or Pg12, the recombinational hypothesis becomes the most likely. For example, in the trait weight of ears, the isolines show a very low value (Table 2), indicating inbreeding depression, ascribed to the homozygosity of the differential chromosome segments linked to the markers. if recombination is assumed in 0000 and 0011 isolines, the resulting genotypes could be those indicated in Table 3a. Recombination in the female parent (0011 --> 001010) could only affect cell (5) of Table 3 because within an isoline there must be genetic variation for the recombination, and in each type of cross a different plant was used as female parent. Recombination in the male parent (0000 --> 000101) must affect the whole column because within an isoline, only one plant was used as male parent. Accepting these two recombinations, the heterotic hypothesis is coherent in all progenies (See Table 3b). Thus, the pale green-12 locus would not be responsible for the differences among genotypes, which should be ascribed to differential loci linked to the marker.
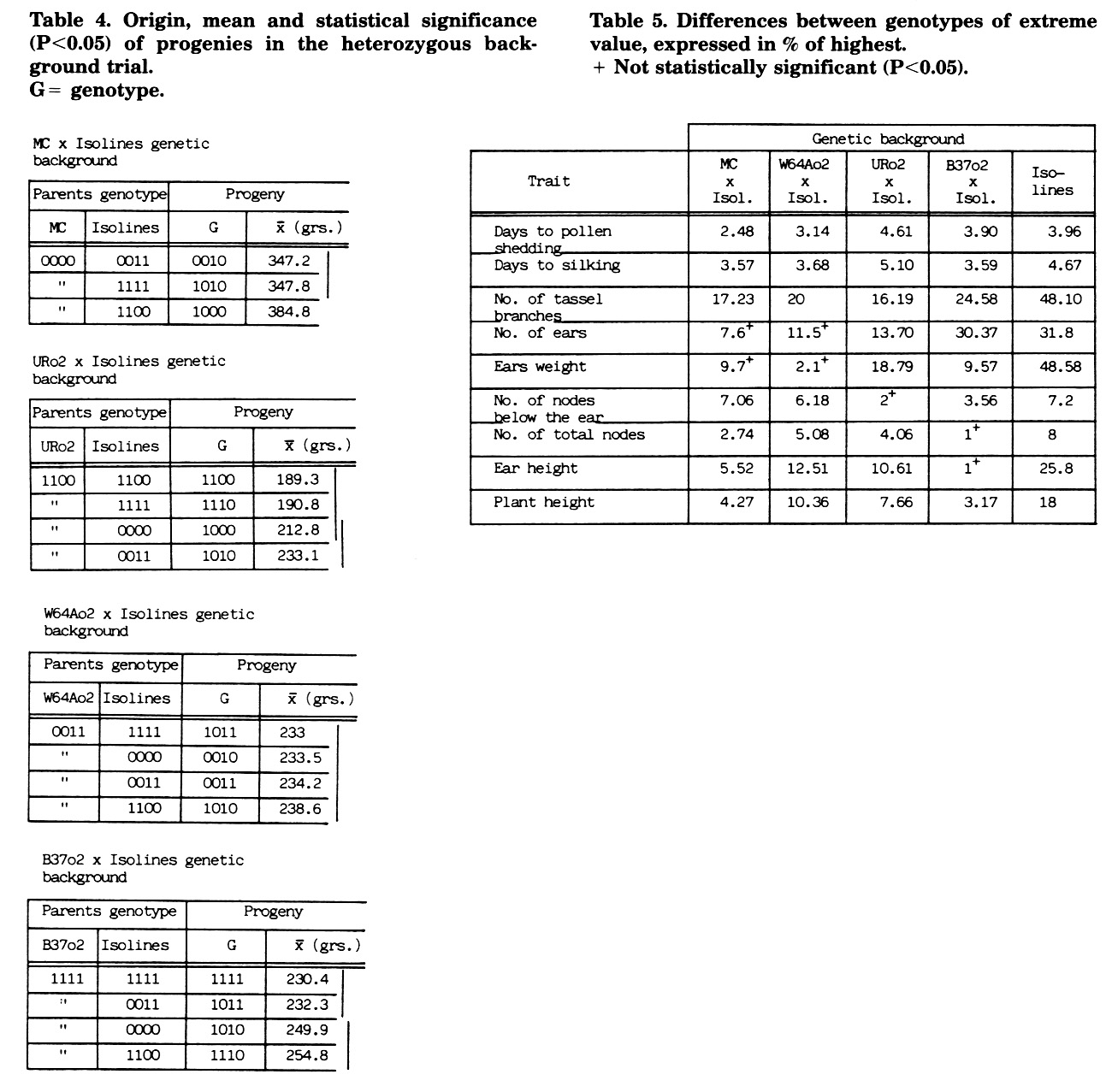
The results in the heterozygous genetic background trial for ear weight are presented in Table 4. In the W64Ao2 x isolines background the double heterozygous genotype 1010 is not significantly distinct from the other genotypes, even from the double homozygous 0011. In the MC x isolines genetic background the double heterozygous genotype is significantly inferior to the 1000 genotype. In the URo2 x isolines genetic background the double heterozygous genotype presents the highest value, easily explained by greater heterozygosity in the differential chromosome segments, due to the origin of isolines from UR x A545o2. We concluded that the distinct "pale green 11, 12" wild genotypes are not different in this character and thus, differences among genotypes within genetic backgrounds are due to loci linked to the markers.
In the other traits, assuming different types of genetic action, analogous results have been obtained. Therefore, the wild genotypes "pale green 11, 12" would not be different in the studied traits.
Differences between genotypes of extreme value, within genetic background, are presented in Table 5. The influence of loci linked to the markers, in all the studied traits, was significant. According to the relative influence of such loci, three groups of traits can be separated: 1) number of tassel branches, number of ears, and weight of ears, 2) upper ear height and plant height, and 3) days to silking and pollen shedding, number of nodes below the ear and number of total nodes.
Table 1. Comparison between two progenies with the same genotype and different origin. * The difference between progenies is significant (P<0.05).
Table 2. Homozygous background trial. a) Origin and value of wild "pale green 11, 12" genotypes. b) Statistical significance (P<0.05). G = genotype.
Table 3. Transformation of Table 2 assuming that the isolines 0000 and 0011 are recombinants. 01 means that the differential segment linked to pg12 has recombinant genes proceeding from the differential segment linked to Pg12. Analogously in 10.
Table 4. Origin, mean and statistical significance (P<0.05) of progenies in the heterozygous background trial. G = genotype.
Table 5. Differences between genotypes of extreme value, expressed in % of highest. + Not statistically significant (P<0.05).
Ll. Bosch, F. Casanas and F. Nuez
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}
{kind=link}