Cms-S can be distinguished from other types of cms on the basis of the two plasmid-like mtDNA's, termed S-1 and S-2 DNAs (Pring et al., P.N.A.S. 74:2904, 1977). Several cytoplasmically reverted strains from male sterility to male fertility have been reported (Singh and Laughnan, Genetics 71:607, 1972), and all of these revertants show the lack of the S-1 and S-2 DNA's (Levings et al., Science 209:1021, 1980).
In this study, we used S-type cytoplasmic lines G, H, J, L, ME, My, R, S, and Vg, which were in WF9 nuclear background. Three lines, J', G', and R' developed from J, G, and R cytoplasms, respectively, produced exceptional male fertile progenies. These revertant lines were advanced crosses with WF9 lacking the nuclear gene Rf3 for fertility restoration of cms-S. In the following generations of crosses with Wf9 (rf3 rf3) males, all of the progenies from the three revertants were male fertile. This suggested that the male fertility was not caused by nuclear mutation of restorer genes, but by cytoplasmically inherited changes.
Mitochondrial DNA (mtDNA) was isolated as described previously (Umbeck and Gengenbach, Crop Sci. 23:584, 1983) from Vg and revertant J', G', and R' cytoplasms possessing WF9 nuclear background. Terminally attached proteins of S-1 and S-2 DNAs were examined by proteinase K treatment prior to phenol-chloroform extraction (Kemble et al., Nucl. Acids Res. 10:8181, 1982). To check S-1 and S-2 DNAs, undigested mtDNAs were analyzed by agarose gel electrophoresis. The restriction sites of chromosomal mtDNA, S-1 and S-2 DNAs were examined by digestion with several restriction enzymes.
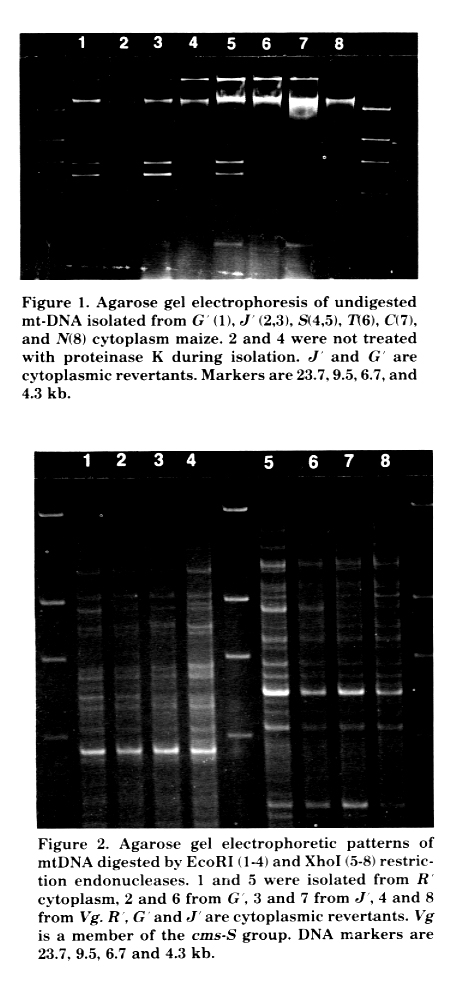
Electrophoresis of undigested mtDNA (Fig. 1) shows that revertants J' and G' have S-1 and S-2 DNAs, and J' has terminally attached proteins like other standard cms-S lines. The restriction fragment patterns in the revertants were different from each other and also differed from that of Vg cytoplasm (Fig. 2). Moreover, these patterns were not identical with those of other revertants reported by Levings et al. The restriction sites of S-1 and S-2 DNA in J', G', R' and Vg cytoplasms were checked by restriction with BamHI, EcoRI, HindIII, PstI, and XhoI (data not given), and there were no restriction site differences detected among these S-1 and S-2 DNAs.
These results indicated that the cytoplasmic changes of J', G' and R' cytoplasm from male sterility to male fertility were associated with rearrangements of mitochondrial chromosomal mtDNA and not with disappearance of plasmid-like S-1 and S-2 DNAs.
Figure 1. Agarose gel electrophoresis of undigested mt-DNA isolated from G'(1), J'(2,3), S(4,5), T(6), C(7), and N(8) cytoplasm maize. 2 and 4 were not treated with proteinase K during isolation. J' and G' are cytoplasmic revertants. Markers are 23.7,9.5,6.7, and 4.3 kb.
Figure 2. Agarose gel electrophoretic patterns of mtDNA digested by EcoRI (1-4) and XhoI (5-8) restriction endonucleases. 1 and 5 were isolated from R' cytoplasm, 2 and 6 from G', 3 and 7 from J', 4 and 8 from Vg. R', G'and J' are cytoplasmic revertants. Vg is a member of the cms-S group. DNA markers are 23.7, 9.5, 6.7 and 4.3 kb.
Teruo Ishige and Burle Gengenbach
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}