In last year's Newsletter we described an unstable bz2 mutable allele (bz2-Mu1) that was isolated from a Mutator line. The original spotted kernel (bz2-Mu1/bz2), when testcrossed and selfed, showed normal segregation, 1:1 and 3:1, respectively. The spotted progeny were again selfed and testcrossed. Although many of the ears yielded the expected segregation ratios, a few plants did not. In the eight plants examined last year, the two plants with a low copy number of Mutator elements and the two plants with approximately five fold more Mutator elements yielded unexpected segregation ratios, while the four plants with intermediate numbers of Mu elements showed normal segregation. This led V. Walbot to postulate that either low or high copy numbers of Mutator elements might inhibit somatic reversion at bz2-Mu1, resulting in fewer spotted kernels than expected. A model was proposed in which lines carrying an intermediate number of Mutator copies would have an active Mutator system.
During the past year, we have continued to analyze whether there is a relationship between Mutator activity and the total copy number of Mutator elements in the genome. We have examined the segregation of Mutator activity in the bz2-Mu1 lineages for two additional generations by following forward mutation at selected loci (presumably measuring transposition) and somatic reversion at bz2-Mu1 (presumably measuring excision at one locus). We have evidence for at least two mechanisms that may control Mutator activity.
We wanted to know whether the somatic reversion at bz2-Mu1 is correlated with Mu activity in general. To do this we have looked at the frequency of seedling mutations (albinos, lethals) in four lineages that vary in Mu copy number and segregation ratios of spotted and bronze kernels. We find that lines with fewer spotted kernels than expected also tend to have a decreased seedling mutation rate. We are therefore using the somatic reversion at bz2-Mu1 as a phenotypic marker for Mutator activity However, this correlation must be made for each lineage examined.
Examination of a much larger sample size than presented last year revealed two plants with equivalent Mu copy number, but one appears to have an active Mutator system, while the other does not. This suggests that the simple model that total copy number controls Mutator activity does not hold for all plants.
We were then interested in testing whether any detectable alteration in the Mu-1 element (as first described by J. Bennetzen) correlated with Mutator activity. DNA from the maize lines described below was digested with the restriction enzyme HinfI and analyzed on Southern blots, probing with the cloned Mu-1 element. HinfI digests within the terminal inverted repeats, resulting in a characteristic 1.3 kbp fragment for Mu-1. Surprisingly, in some of our bz2-Mu1 lines the Mutator elements no longer show the typical restriction pattern of Mu-1, instead many larger fragments are observed. Controls were done to test for completeness of digestion, and these larger fragments are not due to incomplete digestion of total DNA. When the DNA was digested with other enzymes that restrict within the internal portion of the element, fragments larger than those predicted from the restriction map of Mu-1 were also obtained. We are currently investigating whether this altered restriction pattern is due to DNA modification, amplification, or rearrangement of sequences within the element. Results from two lineages are summarized below.
The selfed ear of plant 230-3 (bz2-Mu1/bz2) had no spotted kernels, but behaved normally when testcrossed. Single seedling DNA was prepared from eight kernels off the selfed ear and examined as described above. In all cases no 1.3 kbp HinfI fragments, characteristic of Mu-1, were detectable. Instead, we observed larger Mu-1 homologous fragments ranging in size from approximately 1.4 kbp to 8.0 kbp. We next analyzed both bronze and spotted kernels from the testcrossed ear. Of eight bronze (bz2/bz2) kernels, four contained the characteristic Mu pattern, two contained the 1.3 kbp size class as well as larger fragments, and two contained only larger fragments. Of eight spotted (bz2-Mu1/bz2) kernels, six contained the characteristic Mu pattern, but two contained larger fragments as well as the 1.3 kbp size class. Leaf DNA from the 230-3 plant was analyzed. It contained approximately 30 copies of Mu elements in the characteristic pattern; no larger fragments were observed. This suggests that both altered and unaltered Mu elements can exist together, and that the change can occur within one generation. Two other lineages with aberrant segregation of the mutable phenotype have been analyzed with similar results.
Our working hypothesis is that the altered elements are not functional, and when the element at bz2-Mu1 is altered it is no longer capable of somatic excision. However, not all of our lines that demonstrate aberrant segregation of the mutable phenotype contain this altered pattern. Plant 230-1 (bz2-Mu1/bz2), containing approximately 12 Mu elements, when selfed showed normal segregation, but only 1% spotted kernels were observed in the testcross. Single seedling DNA was prepared from four kernels off each ear. All samples contained only the characteristic Mu HinfI restriction fragments. No larger fragments were observed. The only difference we could detect between the selfed and testcrossed progeny on Southern blots was that the total copy number of Mutator elements tended to be approximately 2-3 fold lower in the backcrossed progeny. Therefore, this plant may be losing Mutator activity due to dilution of Mu copy number or some other alteration not yet detected.
These data suggest there may be at least two mechanisms for decreasing somatic reversion at bz2-Mu1; one, an alteration of the elements as observed in plant 230-3, and two, a dilution of elements as observed for plant 230-1. If the number of functional Mutator elements is too low, somatic excision of the element at bz2-Mu1 may be too infrequent or too late in development to be observed in the aleurone tissue.
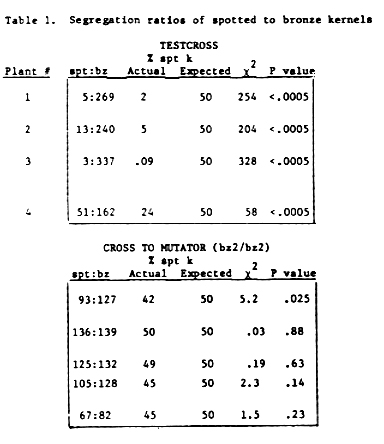
This hypothesis predicts that somatic excision at bz2-Mu1 might be restored by introducing additional elements from an active Mutator line. We have tested whether crossing bz2-Mu1 230-1 progeny to a bz2/bz2 Mutator line would restore somatic reversion at bz2-Mu1. For this experiment kernels from the testcrossed ear were planted and testcrossed to determine which had the bz2-Mu1 allele. (Spotted kernels are observed at low frequency which allows us to determine which plants have the bz2-Mu1 allele.) These plants were also crossed to an active Mutator line (bz2/bz2) to observe somatic reversion at bz2-Mu1. Results are presented in the accompanying table. When the plants were testcrossed, the number of spotted progeny deviated dramatically from that expected for Mendelian segregation, P<.0005. However, when crossed to an active Mutator line, there was an increased number of spotted kernels in all cases, and in 4/5 cases the expected number of spotted progeny were observed. These results suggest that some component in an active Mutator line can stimulate somatic reversion at bz2-Mu1, resulting in normal segregation of the mutable phenotype. Whether this is due to simply increasing the copy number of active Mutator elements, or to the introduction of some other factor, is an open question. Currently, we are investigating whether the seedling mutation rate is also increased in these lines. Similar experiments to determine if the altered elements can be returned to normal by crossing with an active Mutator line are now in progress with progeny from 230-3.
Table 1. Segregation ratios of spotted to bronze kernels
Vicki Chandler and Virginia Walbot
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}