Although maize tissue culture techniques have been improving steadily in recent years, a major limitation still remaining is the poor in vitro response of many agronomically elite genotypes. We conducted a study in 1982 to obtain information on the inheritance of plant regeneration ability of tissue cultures initiated from immature maize embryos. The two populations used in this study were derived from crosses of A188 (excellent tissue culture response) with the inbred lines B73 and A619 (both agronomically elite, but poor in terms of tissue culture response). Each population was composed of six generations (Tables 1 and 2; the reciprocal F1's constitute one generation). Note that F1 embryos are produced on the parental inbreds, and F2 and BC embryos are produced on F1 plants. Immature embryos ranged from 1.0 to 1.5 mm in length. Cultures were scored twice from 14 to 37 days following immature embryo isolation for regenerable structure formation (somatic embryoids, shoot meristems, or scutellar-like bodies).
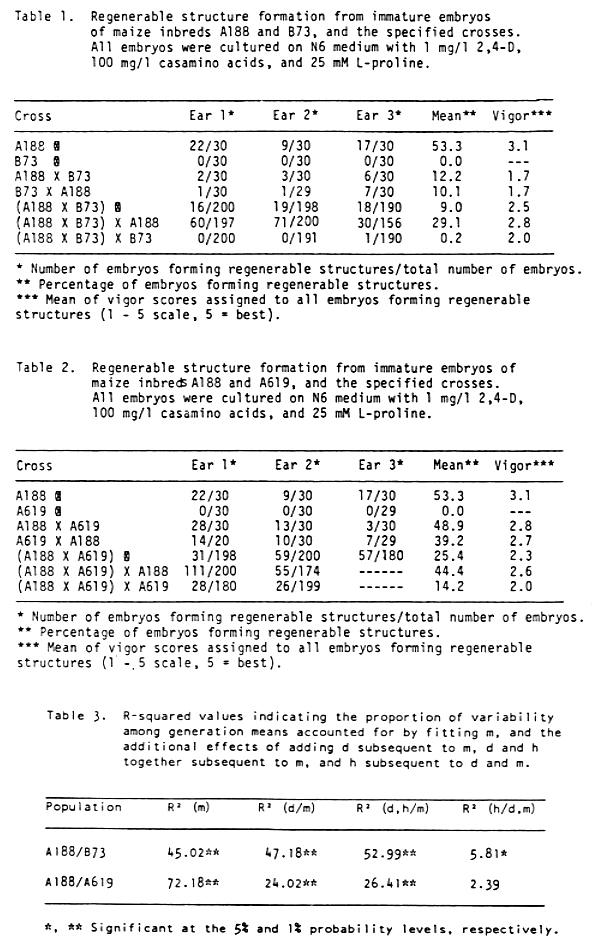
The variation among generations was partitioned using generation means analysis (B. I. Hayman, Heredity 12:371-390, 1958; B. I. Hayman, Genetica 31:133-146, 1960). Least squares regression techniques were used to fit increasingly complex models to generation means of each population (K. Mather and J. L. Jinks, pp. 65-81 in Biometrical Genetics: The Study of Continuous Variation, third edition, Chapman and Hall, N.Y., 1982). The results of the regression analysis were evaluated in terms of the proportion of the uncorrected sums of squares among generation means accounted for by fitting the population mean (m), additive (d), and dominance (h) effects (Table 3).
Mean squares for the pooled additive genetic effects were highly significant for both sets of crosses (Table 3). Selection for regenerable structure formation from maize tissue cultures should therefore be effective within both populations. Mean squares for the additional pooled dominance effects after fitting the pooled additive effects were significant for the A188/B73 crosses, but not for the A188/A619 crosses. The estimate of the parameter h for the pooled dominance effects in the A188/B73 population was negative, indicating that B73 possesses a partially to completely dominant gene or genes which inhibit(s) the formation of regenerable structures. Deviations from the additive and dominance effects were not significant for either population; thus, epistatic effects did not contribute significantly to the variation among generation means. A genetic model based only on the mean and additive genetic effects accounted for 96.2% of the variation among generation means for the A188/A619 population. A model based on the mean and both additive and dominance effects accounted for 98.0% of the variation among the A188/B73 generation means.
In summary, additive genetic effects were highly significant for both the A188/A619 and A188/B73 sets of crosses. Neither dominance nor epistatic effects contributed significantly to the variation among generation means for the A 188/A619 crosses; however, dominance effects were significant for the A188/B73 crosses. The results of this analysis do not rule out the possibility that regenerable structure formation is qualitatively inherited, but we have not yet developed a qualitative model that fits all the data.
Table 1. Regenerable structure formation from immature embryos of maize inbreds A188 and B73, and the specified crosses. All embryos were cultured on N6 medium with 1 mg/l 2,4-D, 100 mg/l casamino acids, and 25 mM L-proline.
Table 2. Regenerable structure formation from immature embryos of maize inbreds A188 and A619, and the specified crosses. All embryos were cultured on N6 medium with 1 mg/l 2,4-D, 100 mg/l casamino acids, and 25 mM L-proline.
Table 3. R-squared values indicating the proportion of variability among generation means accounted for by fitting m, and the additional effects of adding d subsequent to M, d and h together subsequent to m, and h subsequent to d and m.
C. L. Armstrong, C. E. Green, R. L. Phillips and R. E. Stucker
Return to the MNL 59 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}