Excised immature embryos, placed with the scutellum side exposed to air on tissue culture initiation medium B and medium C (see previous article), one week after the initiation of cultures showed proliferating callus tissue at the periphery of the scutellum. Such tissue could be seen in all of the periphery of the scutellum in contact with the medium, and occasionally on the exposed side of the scutellum. However, good callus proliferation was seen on the underside surface of the scutellum. The callus formed was compact, smooth and white (embryogenic callus tissue). Friable and nodular callus (organogenic callus tissue), as obtained in medium A, was not observed in medium B and medium C. Figures 1-6 show embryogenic events in these studies.
After 10 to 15 days in culture, structures resembling coleoptiles were observed on the underside surface of the scutellum. After 3 weeks, such structures appeared as well organized somatic embryos always showing bilateral symmetry and bipolar structure. As reported by Lu et al. (TAG, 62:109-112, 1982), somatic embryos were formed on the same 2,4-D containing media in which the primary culture was obtained. The level of 2,4-D in the medium did not appear to be involved with the embryoid formation over a large range. Both in media with a high level of 2,4-D (3 mg/liter) and in media with a low level (0.5 mg/liter) we observed the same results. However, both the level of sucrose and the high concentration of proline had a strong effect on embryoid formation and on the callus external aspect. We occasionally observed somatic embryogenesis both in media with a normal level of sucrose (2%) and 400 mg proline/liter and in media with a high level of sucrose (12%) without proline. However, only when high levels of both were put together in the medium did we observe a high frequency of somatic embryogenesis. We do not know the reason for such differential response of the embryos.
Sometimes several embryos germinated. However, germinated embryos also produced embryogenic callus. The embryo germination could be due to the fact that several embryos were larger than 2 mm at the time of culture.
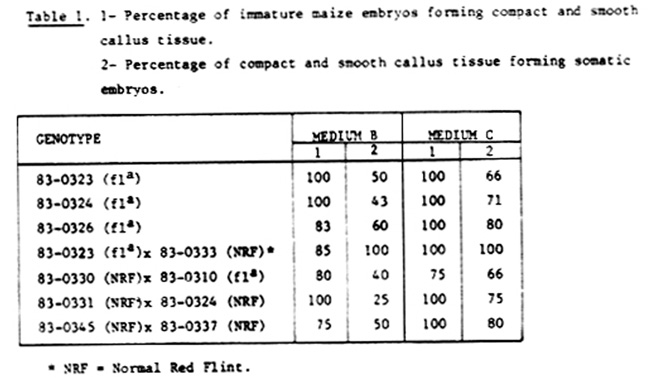
We have detected differences in the percentage of formation of compact and smooth callus tissue and of such callus forming somatic embryos due to the genotype and the effect of the medium (Table 1).
There are great differences in the macro, micro and organic formulation between the MS modified (8) and Yu-pei medium (C). The most important differences are the level Of N03NH4, which is 10 times higher in MS than in Yu-pei medium, and the addition of casein hydrolysate to the Yu-pei medium. Only KN03, KH2PO4, thiamine-HCl and pyridoxine-HCl are more concentrated in the Yu-pei medium than in the MS modified medium. In addition, MoO4Na2, CoCl2, L-asparagine and Ca pantothenate are absent in Yu-pei medium.
As shown in Table 1, we have obtained higher levels of somatic embryogenesis in Yu-pei medium than in MS modified medium. So it would be important to study the effect of some media components on the induction of somatic embryogenesis in maize.
As we have pointed out (Rapela, MNL 57:57-58, 1983), there is also a close relationship between genotype and response to form embryos in culture. The genotype of the donor plant influences both the rate of callus proliferation and somatic embryogenesis induction.
Somatic embryos formed in MS modified medium B and in Yu-pei medium C were transferred to MS medium A without 2,4-D but with 2% sucrose, to induce germination. At the time of writing this communication these embryoids germinated normally.
In our cultures of maize immature embryos on Yu-pei medium C we have observed evidence of direct (nonzygotic) embryogenesis as reported by Conger et al. from mesophyll cells of Dactylis glomerata (Science 221:850-851, 1983). Embryos longer than 2 mm at the time of culture germinated, but such embryos produced embryogenic callus in either medium B or C. Two to 3 weeks before culture initiation the coleoptile of the germinated embryos was 2 to 3 cm long, while the underside surface of the scutellum exhibited embryogenic calli. All the germinated/embryogenic embryos cultured in Yu-pei medium C also showed a probable direct embryo formation on the coleoptile surface. Direct embryo formation began nearer the scutellar node and decreased up to 1 cm from the first point. We have observed between 5 to 15 structures resembling direct embryoids on the surface of each coleoptile that emerged from germinated embryos on medium C. The embryo length ranged from 0.5 to 5 mm and was independent of the position lengthwise on the coleoptile.
Miguel Angel Rapela
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}
{kind=link}