Restoration of the cms-C group of cytoplasms has most recently been shown to be under the control of a single dominant gene, designated Rf4 (Kheyr-Pour et al., Genetics 98:379-388, 1981), although previous work indicated the presence of three or more Rf factors, designated Rf4, Rf5 and Rf6 (Josephson et al., Proc. Ann. Corn Sorghum Res. Conf. 33:48-59, 1978). The fertility response is sporophytic in nature, with all pollen grains from a heterozygote being fully normal.
A series of crosses was made using two different cms-C fertile lines (A619 and NY821LERf), and a series of wx translocation stocks which had been converted to cms-C sterility to detect any possible linkage between the Rf4 locus and a particular wx translocation.
It was hoped that the restorer locus could be mapped to a single chromosome, further reinforcing the recent evidence that a single gene controls this trait.
The series of wx translocations listed in Table 1 was obtained from the Cooperation seed stock and crossed onto an Oh51A cms-C sterile line. Semi-sterile ears indicating presence of the translocation were planted, selfed and the progeny selected for both semi-sterility and segregation of the wx gene. The wx kernels were planted, and sterile plants were paircrossed onto three different cms-C cytoplasm lines:
1) C x A619 (Rf4 Rf4)
2) C x NY821LERf (Rf4 Rf4)
3) C x NYD410 rf4 rf4)
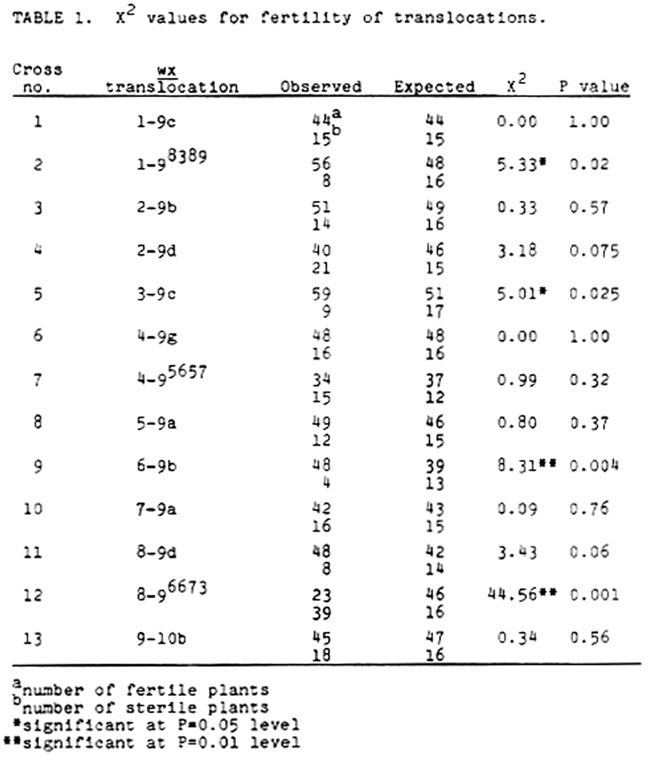
Rows of the NYD410 crosses were planted adjacent to the paired A619 or NY821LERf cross as a check on the cms-C sterility of the wx translocation stock. If the NYD410 paircross was fertile, that wx translocation paircross was discarded. The remaining wx translocation paircrosses were selfed, and only the wx kernels from semi-sterile ears were planted. The plants were scored for sterility of the tassels, with the few occurring late breaks included in the sterile category as this is a recurring problem in cms-C steriles. Any cross exhibiting >25% sterility (nonlinkage gives a 3:1 fertile:sterile ratio) was indicative of linkage of the Rf4 gene to that particular translocation since the wx T stocks were rf4 rf4. Table 1 indicates the results of these crosses and the associated P values assuming a 3:1 ratio.
There are four crosses which differ significantly (P = 0.05 and P = 0.01) from a 3:1 ratio. Three of them (Nos. 2, 5 and 9), which exhibit a much higher degree of fertility than expected, can be explained by misclassification of wx kernels or small sample size. The cross of interest is #12, T8-9(6673), which has a much higher proportion of steriles, suggesting that Rf4 is on chromosome 8. The other translocation involving chromosome 8, T8-9d, shows no evidence of linkage, so the Rf4 locus must be located some distance away from the breakpoint in T8-9d. The reported breakpoints of the two translocations are:
T8-9d: 8L.09 9S.16
T8-9(6673): 8L.35 9S.31
Therefore, the Rf4 locus appears to be on the long arm of chromosome 8, apparently near the end of the chromosome since there is no linkage to the wx gene evident near the 8L.09 breakpoint translocation, and the number of fertiles in the T8-9(6673) cross precludes a tight linkage at the 8L.35 breakpoint, which would be typified by very few fertile plants.
Additional studies to confirm the location of Rf4 on the long arm of chromosome 8 are needed.
TABLE 1. X2 values for fertility of translocations.
Alice Johnson
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}