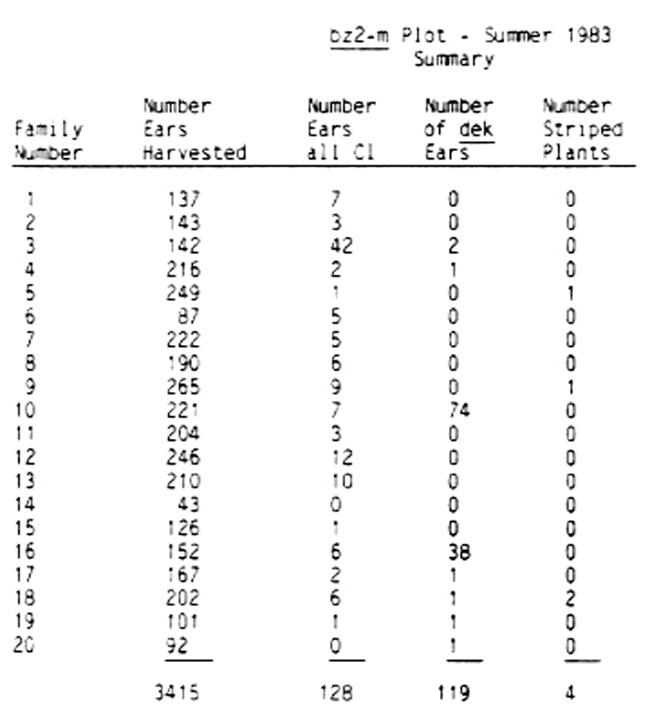
It would be desirable to have a collection of Ds induced defective kernel (dek) mutants since such a collection could aid in determining the nature of the defects in this type of mutant. In particular, such mutants would allow for comparison of the stable mutant condition (in the absence of Ac) with the unstable condition (in the presence of Ac), which should result in the production of sectors of both normal and mutant tissue in the endosperm and embryo of developing kernels. An examination of the developmental fate of such kernels when the size of the genetically normal tissue sectors is varied (by varying the dosage of Ac) may aid in understanding whether or not the defective condition can be rescued by cross- feeding from normal cells in the same tissue, and whether or not plants consisting of normal and mutant sectors can be grown from such kernels. In order to pursue this goal, a stock homozygous for bz2-m and a purple kernel stock homozygous for the normal allele at all of the loci controlling anthocyanin synthesis were planted in the spring of 1982. The stocks and advice were kindly provided by M. G. Neuffer. The kernels of the bz2-m stock selected for planting all carried one dose of Ac at an unlocated site on chromosome 9, as well as Ds at the bz2-m locus. The resulting plants bore ears that produced embryo sacs, half of which contained one dose of Ac and the other half lacked Ac. These ears were pollinated with pollen from the purple stock, resulting in the production of fully colored ears. The kernels on these ears were homozygous for the normal allele at each of the loci involving anthocyanin synthesis, except for being heterozygous at the bronze-2 locus (Bz2/bz2-m). From 20 such ears, 5,906 kernels were obtained and planted as 20 families in the spring of 1983. This planting produced 4,944 plants, from which 3,415 self-pollinated ears were obtained. An average of 195 ears was obtained from the 20 families, and the range of ears per family was from 87 to 265 (see Table). Among the 20 families, 8 yielded one or more ears segregating for dek mutants.
In family #10 there were 141 ears not segregating for dek kernels and 74 ears segregating; in family #16 there were 114 non-segregating ears and 38 segregating ears. In both of these cases it is most likely that the normal allele at a dek locus mutated to the recessive state after zygote formation, but early during the development of the embryo that grew into the bz2-m stock plant used as the female parent in 1982. Such an event could give rise to an ear bearing a large sector (approximately half the ear) that would be +/dek, the other portion being +/+. Kernels from the mutant sector would produce an equal proportion of +/dek and +/+ kernels when pollinated by the purple kernel stock. All of the kernels produced on the non-mutant sector would produce +/+ kernels. For family #3 there were two ears out of 142 harvested that were segregating for dek mutant kernels, and in five other families (#4, 17, 18, 19, and 20) there was one ear segregating for a dek mutant among a large number of ears in each family. In these seven cases the mutation events resulting in the production of dek alleles must have occurred late in ear development of the bz2-m plants used as the female parents in 1982. These events most likely occurred after kernel primordia were formed, but not much later than zygote formation. This appears likely because only one (or 2 in the case of family #3) of the resulting kernels produced a plant carrying the mutant dek allele (as revealed by subsequent self-pollination).
It is not known whether any or all of the new dek mutants were produced by the insertion of a Ds element in a dek locus. It is important in evaluating these data to keep in mind that although the kernels planted in the spring of 1983 were heterozygous at the bz2-m locus (barring earlier changes as proposed to explain the results obtained with families #10 and #16), only one-half of such kernels would contain a single dose of Ac while the other half would lack Ac. Consequently, only that half of the total number of plants which carry Ac could be expected to express mutability in a new dek mutant as well as at the bz2 locus. Some transposition events would most likely result in self-pollinated ears with only a sector of dek kernels rather than a fully segregating ear. Although a detailed scrutiny remains to be done, the preliminary examination of all of the 3,415 ears described above has not identified any sectored ears.
Among the 4,944 plants, there were four plants identified as possible dominant mutants. These were found in three families (#5, #9, and #18) and all four have the same phenotype. They had yellow and green striped leaves either on the top three or four leaves of the main stem (two cases), or on the tillers only (two cases). Future tests are planned to determine the mutability of the new dek mutants, their allelism, and whether they are responsive to Ac. The yellow and green striped plants were outcrossed onto early stocks and the resulting progeny will be examined for this trait.
W. F. Sheridan
Return to the MNL 58 On-Line Index
Return to the Maize Newsletter Index
Return to the MaizeGDB Homepage
{kind=link}